
植物学报 ›› 2019, Vol. 54 ›› Issue (1): 9-22.DOI: 10.11983/CBB18147 cstr: 32102.14.CBB18147
马朝峰( ),戴思兰()
),戴思兰()
收稿日期:2018-06-25
接受日期:2018-09-17
出版日期:2019-01-01
发布日期:2019-07-31
通讯作者:
马朝峰,戴思兰
基金资助:
Chaofeng Ma(),Silan Dai()
Received:2018-06-25
Accepted:2018-09-17
Online:2019-01-01
Published:2019-07-31
Contact:
Chaofeng Ma,Silan Dai
摘要: 光照是影响植物生长发育的重要环境因子, 开花是高等植物生活史上最重要的事件。植物通过光受体感知外界环境中的光照变化, 激活一系列信号转导过程从而适时开花。该文介绍了高等植物光受体的种类、结构特征和生理功能的研究进展, 并系统阐述了红光/远红光受体光敏色素、蓝光受体隐花色素以及FKF1/ZTL/LKP2等介导光信号调控植物开花的分子机制, 包括光受体对CO转录及转录后水平调控和对FT转录水平的调控等。此外, 还介绍了光受体整合光信号与温度和赤霉素等信号调控植物开花的研究进展, 并展望了未来的研究方向。
马朝峰,戴思兰. 光受体介导信号转导调控植物开花研究进展. 植物学报, 2019, 54(1): 9-22.
Chaofeng Ma,Silan Dai. Advances in Photoreceptor-mediated Signaling Transduction in Flowering Time Regulation. Chinese Bulletin of Botany, 2019, 54(1): 9-22.
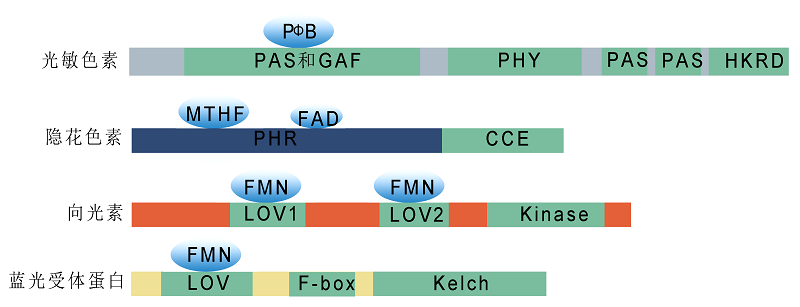
图1 植物光受体结构示意图 光敏色素N端为可共价结合生色团的光感受结构域, 包括PAS、GAF和PHY等亚功能域; C端为光调节结构域, 主要参与光敏色素二聚体的形成及下游的信号转导过程, 包括PAS和HKRD两个亚功能域。隐花色素N端为非共价结合生色团黄素(FAD)和叶酸(MTHF)的光裂解酶相关PHR结构域, C端是对蛋白互作和信号转导十分重要的CCE结构域。向光素通过2个N端的LOV结构域(LOV1和LOV2)结合生色团黄素单核苷酸FMN来感知光照信号, 并通过LOV结构域与辅因子结合或与其它蛋白发生互作。蓝光受体蛋白ZTLs含有3个重要的功能保守结构域: N端的LOV结构域、中间的F-box基序和C端的Kelch重复序列。ZTLs以LOV结构域结合生色团黄素单核苷酸FMN, Kelch结构域则与蛋白质的相互作用相关。
Figure 1 Schematic structural diagrams of plant photoreceptors The N-terminus of phytochromes is a photosensory region (containing PAS, GAF, and PHY), which covalently binds chromophores. The C-terminal light regulatory domains, including PAS and HKRD, are involved in phytochrome dimer formation and downstream signaling transduction. The N-terminus of cryptochromes is a PHR domain, which non-covalently binds FAD and MTHF, and the C-terminal CCE domain is important for protein-protein interactions and signaling transduction. The two N-terminal LOV domains (LOV1 and LOV2) of phototropins bind FMN to perceive light signals and are required for interaction with other proteins. ZTL proteins contain three important functional conserved domains: the N-terminal LOV domain, the intermediate F-box motif, and the C-terminal Kelch repeat region. The LOV domain binds to FMN, and the Kelch domain mediates protein-protein interactions.
| 光受体 | 基因 | 生理功能 | 主要参考文献 |
|---|---|---|---|
| 光敏色素 | PHYA | 促进开花, 幼苗去黄化, 种子萌发, 避阴反应 | Johnson et al., 1994; Sch?fer and Bowler, 2002; Tepperman et al., 2006; Heschel et al., 2007 |
| PHYB | 抑制开花, 幼苗去黄化, 种子萌发, 气孔发育, 避阴反应 | Guo et al., 1998; Sch?fer and Bowler, 2002; Heschel et al., 2007; Wang et al., 2010; Kami et al., 2010 | |
| PHYC | 短日照下抑制开花, 长日照下促进开花 | Monte et al., 2003 | |
| PHYD | 抑制开花, 种子萌发, 避阴反应 | Devlin and Kay, 2000; Sch?fer and Bowler, 2002; Heschel et al., 2007 | |
| PHYE | 抑制开花, 种子萌发, 避阴反应 | Devlin and Kay, 2000; Sch?fer and Bowler, 2002; Heschel et al., 2007 | |
| 隐花色素 | CRY1 | 促进开花, 气孔发育, 幼苗去黄化 | Ahmad and Cashmore, 1993; Mao et al., 2005 |
| CRY2 | 促进开花, 幼苗去黄化, 昼夜节律调控 | Koornneef et al., 1991; Guo et al., 1998 | |
| 向光素 | PHOT1 | 向光性, 光诱导的气孔开放和叶绿体运动, 抑制下胚轴伸长 | Briggs and Christie, 2002; Celaya and Liscum, 2005 |
| PHOT2 | 向光性, 光诱导的气孔开放和叶绿体运动, 抑制下胚轴伸长 | Briggs and Christie, 2002; Celaya and Liscum, 2005 | |
| ZTLs | FKF1 | 促进开花, 昼夜节律信号输入 | Imaizumi et al., 2005; Baudry et al., 2010 |
| ZTL | 长日照下抑制开花, 昼夜节律信号输入 | Somers et al., 2000; Baudry et al., 2010 | |
| LKP2 | 抑制开花 | Takase et al., 2011 | |
| 紫外受体 | UVR8 | 抑制下胚轴伸长, 光形态建成, 抑制开花 | Favory et al., 2009; Gruber et al., 2010; Yang et al., 2018; Liang et al., 2018 |
表1 拟南芥光受体类型及生理功能
Table 1 Photoreceptors and their physiological functions in Arabidopsis thaliana
| 光受体 | 基因 | 生理功能 | 主要参考文献 |
|---|---|---|---|
| 光敏色素 | PHYA | 促进开花, 幼苗去黄化, 种子萌发, 避阴反应 | Johnson et al., 1994; Sch?fer and Bowler, 2002; Tepperman et al., 2006; Heschel et al., 2007 |
| PHYB | 抑制开花, 幼苗去黄化, 种子萌发, 气孔发育, 避阴反应 | Guo et al., 1998; Sch?fer and Bowler, 2002; Heschel et al., 2007; Wang et al., 2010; Kami et al., 2010 | |
| PHYC | 短日照下抑制开花, 长日照下促进开花 | Monte et al., 2003 | |
| PHYD | 抑制开花, 种子萌发, 避阴反应 | Devlin and Kay, 2000; Sch?fer and Bowler, 2002; Heschel et al., 2007 | |
| PHYE | 抑制开花, 种子萌发, 避阴反应 | Devlin and Kay, 2000; Sch?fer and Bowler, 2002; Heschel et al., 2007 | |
| 隐花色素 | CRY1 | 促进开花, 气孔发育, 幼苗去黄化 | Ahmad and Cashmore, 1993; Mao et al., 2005 |
| CRY2 | 促进开花, 幼苗去黄化, 昼夜节律调控 | Koornneef et al., 1991; Guo et al., 1998 | |
| 向光素 | PHOT1 | 向光性, 光诱导的气孔开放和叶绿体运动, 抑制下胚轴伸长 | Briggs and Christie, 2002; Celaya and Liscum, 2005 |
| PHOT2 | 向光性, 光诱导的气孔开放和叶绿体运动, 抑制下胚轴伸长 | Briggs and Christie, 2002; Celaya and Liscum, 2005 | |
| ZTLs | FKF1 | 促进开花, 昼夜节律信号输入 | Imaizumi et al., 2005; Baudry et al., 2010 |
| ZTL | 长日照下抑制开花, 昼夜节律信号输入 | Somers et al., 2000; Baudry et al., 2010 | |
| LKP2 | 抑制开花 | Takase et al., 2011 | |
| 紫外受体 | UVR8 | 抑制下胚轴伸长, 光形态建成, 抑制开花 | Favory et al., 2009; Gruber et al., 2010; Yang et al., 2018; Liang et al., 2018 |
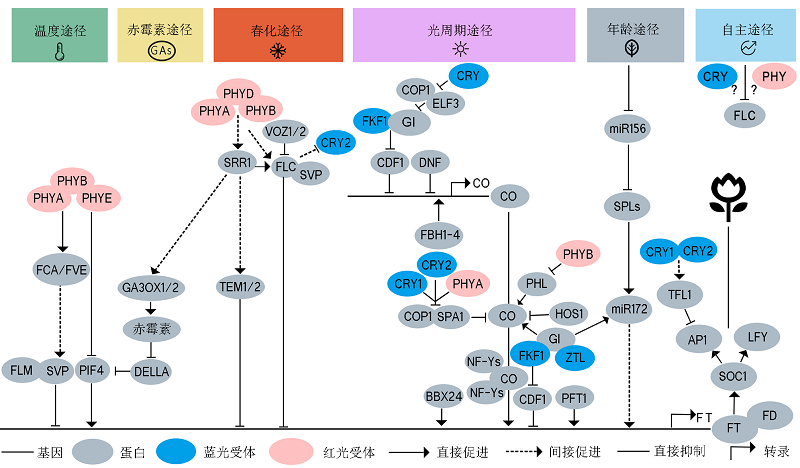
图2 光受体介导环境信号调控拟南芥开花示意图 植物中已探明至少有6种开花调控途径: 光周期途径、春化途径、温度途径、自主途径、赤霉素途径和年龄途径。在叶片中, 光敏色素、隐花色素和ZTL/FKF1/LKP2等光受体介导光信号并将信号传递给昼夜节律钟, 经过多种开花途径的信号整合, 最终直接或间接调控CO、FT和FLC的表达和蛋白稳定性。温度途径中光敏色素通过FCA/FVE间接调控FLM、SVP和PIF4进而调控FT的转录; 赤霉素途径和春化途径由SRR1进行信号整合, 通过GA生物合成途径GA3OX1/2以及DELLA蛋白调控FT的转录, 春化途径通过VOZ1/2和光敏色素调控FLC、SVP和TEM1/2进而调控FT的表达; 自主途径通过抑制开花阻遏物和春化途径关键基因FLC促进开花; 光受体在光周期途径中发挥重要作用, GI、CDF1、DNF、FBH1-4、COP1、SPA1、HOS1和NF-Ys等蛋白直接或间接与光受体互作, 对CO转录、转录后水平以及FT的转录水平等关键调控节点进行调控。年龄途径通过miR156/172和SPLs调控FT的表达。不同的开花途径之间存在信号整合机制, 最终开花信号被整合到FT、SOC1和LFY, 激活AP1和LFY, 进而完成开花起始。实线箭头表示直接促进, 虚线箭头表示间接促进, 钝化线表示直接抑制, 红色表示红光/远红光受体, 蓝色表示蓝光受体, 灰色表示信号蛋白。
Figure 2 A schematic diagram of flowering time regulation by photoreceptor-mediated environmental signaling in Arabidopsis Six flowering regulatory pathways have been identified in plants, including photoperiodic pathway, vernalization pathway, temperature pathway, autonomous pathway, gibberellin pathway, and age pathway. Phytochromes, cryptochromes, and ZTL/FKF1/ LKP2 perceive light signals in leaves, and transmit the signals to the circadian clock. After signaling integration through multiple flowering pathways, the photoreceptors eventually directly or indirectly regulate CO, FT and FLC expression and protein stability. In the ambient temperature pathway, the phytochromes indirectly regulate the transcription of FT by FLM, SVP and PIF4 through FCA/FVE. SRR1 integrates signals from the gibberellin pathway and vernalization pathway and regulate FT transcription through GA3OX1/2 and DELLA proteins in the GA pathway. In the vernalization pathway, VOZ1/2 and phytochromes regulates FT expression through FLC, SVP, and TEM1/2. The autonomous pathway promotes flowering by repressing FLC. Photoreceptors play an important role in the photoperiodic pathway. GI, CDF1, DNF, FBH1-4, COP1, SPA1, HOS1, and NF-Ys directly or indirectly interact with photoreceptors to regulate CO transcription, CO stability, and FT transcription. The age pathway regulates FT expression through miR156/172 and SPLs. There are signal integration mechanisms between different flowering pathways. The signals are integrated into FT, SOC1, and LFY, leading to the activation of AP1 and LFY and eventually the initiation of flowering. Solid arrows indicate direct promotion; dotted arrows indicate indirect promotion; blunted lines indicate direct inhibition; red ovals indicate red/far-red light photoreceptors; blue ovals indicate blue light photoreceptors, and gray ovals indicates signaling intermediate proteins.
| 37 |
Halliday KJ, Salter MG, Thingnaes E, Whitelam GC ( 2003). Phytochrome control of flowering is temperature sensitive and correlates with expression of the floral integrator FT. Plant J 33, 875-885.
DOI URL PMID |
| 38 |
Helliwell CA, Wood CC, Robertson M, Peacock WJ, Dennis ES ( 2006). The Arabidopsis FLC protein interacts directly in vivo with SOC1 and FT chromatin and is part of a high-molecular-weight protein complex.Plant J 46, 183-192.
DOI URL PMID |
| 39 |
Heschel MS, Selby J, Butler C, Whitelam GC, Sharrock RA, Donohue K ( 2007). A new role for phytochromes in temperature-dependent germination. New Phytol 174, 735-741.
DOI URL PMID |
| 40 |
Huq E, Tepperman JM, Quail PH ( 2000). GIGANTEA is a nuclear protein involved in phytochrome signaling in Ara- bidopsis. Proc Natl Acad Sci USA 97, 9789-9794.
DOI URL PMID |
| 41 | Imaizumi T, Schultz TF, Harmon FG, Ho LA, Kay SA ( 2005). FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis.Science 309, 293-297. |
| 42 | Iñigo SS, Alvarez MJ, Strasser B, Califano A, Cerdán PD ( 2012). PFT1, the MED25 subunit of the plant mediator complex, promotes flowering through CONSTANS depen- dent and independent mechanisms in Arabidopsis. Plant J 69, 601-612. |
| 43 |
Ito S, Song YH, Imaizumi T ( 2012 a). LOV domain-contai- ning F-box proteins: light-dependent protein degradation modules in Arabidopsis. Mol Plant 5, 573-582.
DOI URL PMID |
| 44 |
Ito S, Song YH, Josephson-Day AR, Miller RJ, Breton G, Olmstead RG, Imaizumi T ( 2012 b). Flowering BHLH transcriptional activators control expression of the photoperiodic flowering regulator CONSTANS in Arabidopsis . Proc Natl Acad Sci USA 109, 3582-3587.
DOI URL PMID |
| 45 | Jang S, Marchal V, Panigrahi KCS, Wenkel S, Soppe W, Deng XW, Valverde F, Coupland G ( 2008). Arabidopsis COP1 shapes the temporal pattern of CO accumulation conferring a photoperiodic flowering response. EMBO J 27, 1277-1288. |
| 46 |
Jarillo JA, Capel J, Tang RH, Yang HQ, Alonso JM, Ecker JR, Cashmore AR ( 2001). An Arabidopsis circadian clock component interacts with both CRY1 and phyB. Nature 410, 487-490.
DOI URL PMID |
| 47 |
Jenkins GI ( 2014). The UV-B photoreceptor UVR8: from structure to physiology. Plant Cell 26, 21-37.
DOI URL PMID |
| 48 | Johansson M, Staiger D ( 2014). SRR1 is essential to repress flowering in non-inductive conditions in Arabidopsis thaliana.J Exp Bot 65, 5811-5822. |
| 49 |
Johansson M, Staiger D ( 2015). Time to flower: interplay between photoperiod and the circadian clock. J Exp Bot 66, 719-730.
DOI URL PMID |
| 50 |
Johnson E, Bradley M, Harberd NP, Whitelam GC ( 1994). Photoresponses of light-grown phyA mutants of Arabidopsis (Phytochrome A is required for the perception of daylength extensions).Plant Physiol 105, 141-149.
DOI URL PMID |
| 51 |
Kami C, Lorrain S, Hornitschek P, Fankhauser C ( 2010). Chapter two—light-regulated plant growth and development. Curr Top Dev Biol 91, 29-66.
DOI URL |
| 52 |
Kim DH, Doyle MR, Sung SB, Amasino RM ( 2009). Vernalization: winter and the timing of flowering in plants. Annu Rev Cell Dev Biol 25, 277-299.
DOI URL PMID |
| 53 |
Kim J, Geng RS, Gallenstein RA, Somers DE ( 2013). The F-box protein ZEITLUPE controls stability and nucleocytoplasmic partitioning of GIGANTEA. Development 140, 4060-4069.
DOI URL PMID |
| 54 | Koornneef M, Hanhart CJ, van der Veen JH ( 1991). A genetic and physiological analysis of late flowering mutants in Arabidopsis thaliana.Mol Gen Genet 229, 57-66. |
| 55 |
Kumar SV, Lucyshyn D, Jaeger KE, Alós E, Alvey E, Harberd NP, Wigge PA ( 2012). Transcription factor PIF4 controls the thermosensory activation of flowering. Nature 484, 242-245.
DOI URL PMID |
| 56 |
Kumimoto RW, Adam L, Hymus GJ, Repetti PP, Reuber TL, Marion CM, Hempel FD, Ratcliffe OJ ( 2008). The Nuclear Factor Y subunits NF-YB2 and NF-YB3 play additive roles in the promotion of flowering by inductive long- day photoperiods in Arabidopsis. Planta 228, 709-723.
DOI URL PMID |
| 57 |
Laubinger S, Marchal V, Gentilhomme J, Wenkel S, Adrian J, Jang S, Kulajta C, Braun H, Coupland G, Hoecker U ( 2006). Arabidopsis SPA proteins regulate photoperiodic flowering and interact with the floral inducer CONSTANS to regulate its stability. Development 133, 3213-3222.
DOI URL PMID |
| 58 |
Lazaro A, Valverde F, Piñeiro M, Jarillo JA ( 2012). The Arabidopsis E3 ubiquitin ligase HOS1 negatively regulates CONSTANS abundance in the photoperiodic control of flowering. Plant Cell 24, 982-999.
DOI URL PMID |
| 59 |
Lee JH, Yoo SJ, Park SH, Hwang I, Lee JS, Ahn JH ( 2007). Role of SVP in the control of flowering time by ambient temperature in Arabidopsis.Genes Dev 21, 397-402.
DOI URL PMID |
| 60 |
Leivar P, Monte E ( 2014). PIFs: systems integrators in plant development. Plant Cell 26, 56-78.
DOI URL PMID |
| 61 | Li F, Sun JJ, Wang DH, Bai SN, Clarke AK, Holm M ( 2014). The B-Box Family Gene STO (BBX24) in Arabidopsis tha- liana regulates flowering time in different pathways.PLoS One 9, e87544. |
| 62 |
Liang T, Mei SL, Shi C, Yang Y, Peng Y, Ma LB, Wang F, Li X, Huang X, Yin YH, Liu HT ( 2018). UVR8 interacts with BES1 and BIM1 to regulate transcription and photomorphogenesis in Arabidopsis. Dev Cell 44, 512-523.
DOI URL PMID |
| 63 | Liu B, Zuo ZC, Liu HT, Liu XM, Lin CT ( 2011). Arabidopsis cryptochrome 1 interacts with SPA1 to suppress COP1 activity in response to blue light. Genes Dev 25, 1029-1034. |
| 64 | Liu HT, Wang Q, Liu YW, Zhao XY, Imaizumi T, Somers DE, Tobin EM, Lin CT ( 2013). Arabidopsis CRY2 and ZTL mediate blue-light regulation of the transcription factor CIB1 by distinct mechanisms. Proc Natl Acad Sci USA 110, 17582-17587. |
| 65 |
Liu HT, Yu XH, Li KW, Klejnot J, Yang HY, Lisiero D, Lin CT ( 2008). Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis. Science 322, 1535-1539.
DOI URL PMID |
| 66 | Mao J, Zhang YC, Sang Y, Li QH, Yang HQ ( 2005). From the cover: a role for Arabidopsis cryptochromes and COP1 in the regulation of stomatal opening. Proc Natl Acad Sci USA 102, 12270-12275. |
| 1 | 许淑娟, 种康 ( 2018). “先驱”转录因子LEC1在早期胚胎重置春化状态的机制. 植物学报 53, 1-4. |
| 2 | Ahmad M, Cashmore AR ( 1993). HY4 gene of A.thaliana encodes a protein with characteristics of a blue-light photoreceptor..Nature 366, 162-166. |
| 67 | Más P, Kim WY, Somers DE, Kay SA ( 2003). Targeted degradation of TOC1 by ZTL modulates circadian function in Arabidopsis thaliana.Nature 426, 567-570. |
| 68 |
Matsushita T, Mochizuki N, Nagatani A ( 2003). Dimers of the N-terminal domain of phytochrome B are functional in the nucleus. Nature 424, 571-574.
DOI URL PMID |
| 69 |
Mizoguchi T, Wright L, Fujiwara S, Cremer F, Lee K, Onouchi H, Mouradov A, Fowler S, Kamada H, Putterill J, Coupland G ( 2005). Distinct roles of GIGANTEA in promoting flowering and regulating circadian rhythms in Arabidopsis.Plant Cell 17, 2255-2270.
DOI URL PMID |
| 70 | Monte E, Alonso JM, Ecker JR, Zhang YL, Li X, Young J, Austin-Phillips S, Quail PH ( 2003). Isolation and characterization of phyC mutants in Arabidopsis reveals complex crosstalk between phytochrome signaling pathways.Plant Cell 15, 1962-1980. |
| 3 |
Amasino R ( 2010). Seasonal and developmental timing of flowering. Plant J 61, 1001-1013.
DOI URL PMID |
| 4 | Andrés F, Coupland G ( 2012). The genetic basis of flowe- ring responses to seasonal cues. Nat Rev Genet 13, 627-639. |
| 71 | Morris K, Thornber S, Codrai L, Richardson C, Craig A, Sadanandom A, Thomas B, Jackson S ( 2010). DAY NEUTRAL FLOWERING represses CONSTANS to prevent Arabidopsis flowering early in short days.Plant Cell 22, 1118-1128. |
| 72 |
Nelson DC, Lasswell J, Rogg LE, Cohen MA, Bartel B ( 2000). FKF1, a clock-controlled gene that regulates the transition to flowering in Arabidopsis. Cell 101, 331-340.
DOI URL PMID |
| 73 |
Osnato M, Castillejo C, Matías-Hernández L, Pelaz S ( 2012). TEMPRANILLO genes link photoperiod and gibberellin pathways to control flowering in Arabidopsis. Nat Commun 3, 808.
DOI URL PMID |
| 74 |
Posé D, Verhage L, Ott F, Yant L, Mathieu J, Angenent GC, Immink RGH, Schmid M ( 2013). Temperature- dependent regulation of flowering by antagonistic FLM variants. Nature 503, 414-417.
DOI URL PMID |
| 5 |
Bae G, Choi G ( 2008). Decoding of light signals by plant phytochromes and their interacting proteins. Annu Rev Plant Biol 59, 281-311.
DOI URL PMID |
| 6 | Balasubramanian S, Sureshkumar S, Lempe J, Weigel D ( 2006). Potent induction of Arabidopsis thaliana flowering by elevated growth temperature.PLoS Genet 2, e106. |
| 75 |
Qian CZ, Mao WW, Liu Y, Ren H, Lau OS, Ouyang XH, Huang X ( 2016). Dual-source nuclear monomers of UV-B light receptor direct photomorphogenesis in Arabidopsis. Mol Plant 9, 1671-1674.
DOI URL PMID |
| 76 | Rataj K, Simpson GG ( 2014). Message ends: RNA 3' pro- cessing and flowering time control. J Exp Bot 65, 353-363. |
| 77 | Reinhart BJ, Weinstein EG, Rhoades MW, Bartel B, Bartel DP ( 2002). MicroRNAs in plants. Genes Dev 16, 1616-1626. |
| 78 |
Rizzini L, Favory JJ, Cloix C, Faggionato D, O'Hara A, Kaiserli E, Baumeister R, Schäfer E, Nagy F, Jenkins GI, Ulm R ( 2011). Perception of UV-B by the Arabidopsis UVR8 protein. Science 332, 103-106.
DOI URL PMID |
| 7 | Baudry A, Ito S, Song YH, Strait AA, Kiba T, Lu S, Henriques R, Pruneda-Paz JL, Chua NH, Tobin EM, Kay SA, Imaizumi T ( 2010). F-box proteins FKF1 and LKP2 act in concert with ZEITLUPE to control Arabidopsis clock progression. Plant Cell 22, 606-622. |
| 8 |
Bendix C, Marshall CM, Harmon FG ( 2015). Circadian clock genes universally control key agricultural traits. Mol Plant 8, 1135-1152.
DOI URL PMID |
| 79 |
Rockwell NC, Su YS, Lagarias JC ( 2006). Phytochrome structure and signaling mechanisms. Annu Rev Plant Biol 57, 837-858.
DOI URL PMID |
| 80 | Salathia N, Davis SJ, Lynn JR, Michaels SD, Amasino RM, Millar AJ ( 2006). FLOWERING LOCUS C-dependent and -independent regulation of the circadian clock by the autonomous and vernalization pathways. BMC Plant Biol 6, 10. |
| 9 | Blázquez MA, Ahn JH, Weigel D ( 2003). A thermosensory pathway controlling flowering time in Arabidopsis thaliana.Nat Genet 33, 168-171. |
| 10 |
Blümel M, Dally N, Jung C ( 2015). Flowering time regulation in crops-what did we learn from Arabidopsis? Curr Opin Biotechnol 32, 121-129.
DOI URL PMID |
| 81 |
Samach A, Onouchi H, Gold SE, Ditta GS, Schwarz- Sommer Z, Yanofsky MF, Coupland G ( 2000). Distinct roles of CONSTANS target genes in reproductive deve- lopment of Arabidopsis. Science 288, 1613-1616.
DOI URL PMID |
| 82 |
Samach A, Wigge PA ( 2005). Ambient temperature perception in plants. Curr Opin Plant Biol 8, 483-486.
DOI URL PMID |
| 83 | Sawa M, Kay SA ( 2011). GIGANTEA directly activates Flowering Locus T in Arabidopsis thaliana.Proc Natl Acad Sci USA 108, 11698-11703. |
| 84 | Sawa M, Nusinow DA, Kay SA, Imaizumi T ( 2007). FKF1 and GIGANTEA complex formation is required for day- length measurement in Arabidopsis. Science 318, 261-265. |
| 11 |
Briggs WR, Christie JM ( 2002). Phototropins 1 and 2: versatile plant blue-light receptors. Trends Plant Sci 7, 204-210.
DOI URL PMID |
| 12 |
Brudler R, Hitomi K, Daiyasu H, Toh H, Kucho K, Ishiura M, Kanehisa M, Roberts VA, Todo T, Tainer JA, Getzoff ED ( 2003). Identification of a new cryptochrome class: structure, function, and evolution. Mol Cell 11, 59-67.
DOI URL PMID |
| 85 |
Schäfer E, Bowler C ( 2002). Phytochrome-mediated photoperception and signal transduction in higher plants. EMBO Rep 3, 1042-1048.
DOI URL |
| 86 | Schultz TF, Kiyosue T, Yanovsky M, Wada M, Kay SA ( 2001). A role for LKP2 in the circadian clock of Arabidopsis. Plant Cell 13, 2659-2670. |
| 13 |
Cai XN, Ballif J, Endo S, Davis E, Liang MX, Chen D, DeWald D, Kreps J, Zhu T, Wu YJ ( 2007). A putative CCAAT-binding transcription factor is a regulator of flowe- ring timing in Arabidopsis. Plant Physiol 145, 98-105.
DOI URL PMID |
| 14 | Cao SH, Kumimoto RW, Gnesutta N, Calogero AM, Mantovani R, Holt BF ( 2014). A distal CCAAT/NUCLEAR FACTOR Y complex promotes chromatin looping at the FLOWERING LOCUS T promoter and regulates the timing of flowering in Arabidopsis.Plant Cell 26, 1009-1017. |
| 87 |
Sheerin DJ, Menon C, zur Oven-Krockhaus S, Enderle B, Zhu L, Johnen P, Schleifenbaum F, Stierhof YD, Huq E, Hiltbrunner A ( 2015). Light-activated phytochrome A and B interact with members of the SPA family to promote photomorphogenesis in Arabidopsis by reorganizing the COP1/SPA complex. Plant Cell 27, 189-201.
DOI URL PMID |
| 88 | Somers DE, Kim WY, Geng RS ( 2004). The F-box protein ZEITLUPE confers dosage-dependent control on the circadian clock, photomorphogenesis, and flowering time. Plant Cell 16, 769-782. |
| 89 |
Somers DE, Schultz TF, Milnamow M, Kay SA ( 2000). ZEITLUPE encodes a novel clock-associated PAS protein from Arabidopsis . Cell 101, 319-329.
DOI URL PMID |
| 90 |
Song YH, Smith RW, To BJ, Millar AJ, Imaizumi T ( 2012). FKF1 conveys timing information for CONSTANS stabilization in photoperiodic flowering. Science 336, 1045-1049.
DOI URL PMID |
| 15 |
Cashmore AR ( 2003). Cryptochromes: enabling plants and animals to determine circadian time. Cell 114, 537-543.
DOI URL PMID |
| 16 | Cashmore AR, Jarillo JA, Wu YJ, Liu DM ( 1999). Cryptochromes: blue light receptors for plants and animals. Science 284, 760-765. |
| 17 |
Castillejo C, Pelaz S ( 2008). The balance between CONSTANS and TEMPRANILLO activities determines FT expression to trigger flowering.Curr Biol 18, 1338-1343.
DOI URL PMID |
| 18 | Celaya RB, Liscum E ( 2005). Phototropins and associated signaling: providing the power of movement in higher plants. Photochem Photobiol 81, 73-80. |
| 91 | Staiger D, Allenbach L, Salathia N, Fiechter V, Davis SJ, Millar AJ, Chory J, Fankhauser C ( 2003). The Arabidopsis SRR1 gene mediates phyB signaling and is required for normal circadian clock function.Genes Dev 17, 256-268. |
| 92 |
Steinbach Y, Hennig L ( 2014). Arabidopsis MSI1 functions in photoperiodic flowering time control. Front Plant Sci 5, 77.
DOI URL PMID |
| 19 |
Chaves I, Pokorny R, Byrdin M, Hoang N, Ritz T, Brettel K, Essen LO, van der Horst GTJ, Batschauer A, Ahmad M ( 2011). The cryptochromes: blue light photoreceptors in plants and animals. Annu Rev Plant Biol 62, 335-364.
DOI URL PMID |
| 20 |
Chen M, Chory J ( 2011). Phytochrome signaling mechanisms and the control of plant development. Trends Cell Biol 21, 664-671.
DOI URL PMID |
| 93 |
Strasser B, Alvarez MJ, Califano A, Cerdán PD ( 2009). A complementary role for ELF3 and TFL1 in the regulation of flowering time by ambient temperature. Plant J 58, 629-640.
DOI URL PMID |
| 94 |
Suetsugu N, Wada M ( 2013). Evolution of three LOV blue light receptor families in green plants and photosynthetic stramenopiles: phototropin, ZTL/FKF1/LKP2 and aureochrome. Plant Cell Physiol 54, 8-23.
DOI URL PMID |
| 95 | Takano M, Inagaki N, Xie XZ, Yuzurihara N, Hihara F, Ishizuka T, Yano M, Nishimura M, Miyao A, Hirochika H, Shinomura T ( 2005). Distinct and cooperative functions of phytochromes A, B, and C in the control of deetiolation and flowering in rice. Plant Cell 17, 3311-3325. |
| 96 | Takase T, Nishiyama Y, Tanihigashi H, Ogura Y, Miyazaki Y, Yamada Y, Kiyosue T ( 2011). LOV KELCH PROTEIN2 and ZEITLUPE repress Arabidopsis photoperiodic flowe- ring under non-inductive conditions, dependent on FLAVIN- BINDING KELCH REPEAT F-BOX1.Plant J 67, 608-621. |
| 97 |
Tao Z, Shen LS, Gu XF, Wang YZ, Yu H, He YH ( 2017). Embryonic epigenetic reprogramming by a pioneer transcription factor in plants. Nature 551, 124-128.
DOI URL PMID |
| 98 |
Teotia S, Tang GL ( 2015). To bloom or not to bloom: role of microRNAs in plant flowering. Mol Plant 8, 359-377.
DOI URL PMID |
| 99 |
Tepperman JM, Hwang YS, Quail PH ( 2006). phyA dominates in transduction of red-light signals to rapidly responding genes at the initiation of Arabidopsis seedling de-etiolation. Plant J 48, 728-742.
DOI URL PMID |
| 100 | Tiwari SB, Shen Y, Chang HC, Hou YL, Harris A, Ma SF, McPartland M, Hymus GJ, Adam L, Marion C, Belachew A, Repetti PP, Reuber TL, Ratcliffe OJ ( 2010). The flowering time regulator CONSTANS is recruited to the FLOWERING LOCUS T promoter via a unique cis-ele- ment.New Phytol 187, 57-66. |
| 101 | Valverde F, Mouradov A, Soppe W, Ravenscroft D, Samach A, Coupland G ( 2004). Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science 303, 1003-1006. |
| 102 |
Wang CQ, Guthrie C, Sarmast MK, Dehesh K ( 2014). BBX19 interacts with CONSTANS to repress FLOWE- RING LOCUS T transcription, defining a flowering time checkpoint in Arabidopsis.Plant Cell 26, 3589-3602.
DOI URL PMID |
| 21 |
Chen XM ( 2004). A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development.Science 303, 2022-2025.
DOI URL PMID |
| 22 | Christie JM ( 2007). Phototropin blue-light receptors. Annu Rev Plant Biol 58, 21-45. |
| 103 | Wang FF, Lian HL, Kang CY, Yang HQ ( 2010). Phytochrome B is involved in mediating red light-induced sto- matal opening in Arabidopsis thaliana.Mol Plant 3, 246-259. |
| 104 |
Wang JW ( 2014). Regulation of flowering time by the miR156-mediated age pathway. J Exp Bot 65, 4723-4730.
DOI URL PMID |
| 105 |
Wang Q, Zuo ZC, Wang X, Gu LF, Yoshizumi T, Yang ZH, Yang L, Liu Q, Liu W, Han YJ, Kim JI, Liu B, Wohlschlegel JA, Matsui M, Oka Y, Lin CT ( 2016). Photoactivation and inactivation of Arabidopsis cryptochrome 2. Science 354, 343-347.
DOI URL PMID |
| 106 |
Wu D, Hu Q, Yan Z, Chen W, Yan CY, Huang X, Zhang J, Yang PY, Deng HT, Wang JW, Deng XW, Shi YG ( 2012). Structural basis of ultraviolet-B perception by UVR8. Nature 484, 214-219.
DOI URL PMID |
| 107 |
Wu G, Park MY, Conway SR, Wang JW, Weigel D, Poethig RS ( 2009). The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 138, 750-759.
DOI URL PMID |
| 23 |
Christie JM, Arvai AS, Baxter KJ, Heilmann M, Pratt AJ, O'Hara A, Kelly SM, Hothorn M, Smith BO, Hitomi K, Jenkins GI, Getzoff ED ( 2012). Plant UVR8 photoreceptor senses UV-B by tryptophan-mediated disruption of cross-dimer salt bridges. Science 335, 1492-1496.
DOI URL PMID |
| 24 |
Coelho CP, Minow MAA, Chalfun-Júnior A, Colasanti J ( 2014). Putative sugarcane FT/TFL1 genes delay flowe- ring time and alter reproductive architecture in Arabidopsis.Front Plant Sci 5, 221.
DOI URL PMID |
| 108 | Xing SP, Salinas M, Höhmann S, Berndtgen R, Huijser P ( 2010). miR156-targeted and nontargeted SBP-box transcription factors act in concert to secure male fertility in Arabidopsis. Plant Cell 22, 3935-3950. |
| 109 | Yamaguchi A, Kobayashi Y, Goto K, Abe M, Araki T ( 2005). TWIN SISTER OF FT ( TSF) acts as a floral pathway integrator redundantly with FT.Plant Cell Physiol 46, 1175-1189. |
| 25 |
Corbesier L, Vincent C, Jang S, Fornara F, Fan QZ, Searle I, Giakountis A, Farrona S, Gissot L, Turnbull C, Coupland G ( 2007). FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 316, 1030-1033.
DOI URL PMID |
| 26 | de Lucas M, Davière JM, Rodríguez-Falcón M, Pontin M, Iglesias-Pedraz JM, Lorrain S, Fankhauser C, Blázquez MA, Titarenko E, Prat S ( 2008). A molecular framework for light and gibberellin control of cell elongation. Nature 451, 480-484. |
| 110 |
Yan ZQ, Liang DW, Liu H, Zheng GC ( 2010). FLC: a key regulator of flowering time in Arabidopsis. Russ J Plant Physiol 57, 166-174.
DOI URL |
| 111 |
Yang HQ, Wu YJ, Tang RH, Liu DM, Liu Y, Cashmore AR ( 2000). The C termini of Arabidopsis cryptochromes mediate a constitutive light response. Cell 103, 815-827.
DOI URL PMID |
| 112 | Yang LW, Fu JX, Qi S, Hong Y, Huang H, Dai SL ( 2017). Molecular cloning and function analysis of ClCRY1a and ClCRY1b, two genes in Chrysanthemum lavandulifolium that play vital roles in promoting floral transition.Gene 617, 32-43. |
| 113 |
Yang Y, Liang T, Zhang LB, Shao K, Gu XX, Shang RX, Shi N, Li X, Zhang P, Liu HT ( 2018). UVR8 interacts with WRKY36 to regulate HY5 transcription and hypocotyl elongation in Arabidopsis. Nat Plants 4, 98-107.
DOI URL PMID |
| 114 |
Yasui Y, Kohchi T ( 2014). VASCULAR PLANT ONE-ZINC FINGER1 and VOZ2 repress the FLOWERING LOCUS C clade members to control flowering time in Arabidopsis.Biosci Biotechnol Biochem 78, 1850-1855.
DOI URL |
| 115 | Yasui Y, Mukougawa K, Uemoto M, Yokofuji A, Suzuri R, Nishitani A, Kohchi T ( 2012). The phytochrome-interac-ting VASCULAR PLANT ONE-ZINC FINGER 1 and VOZ2 redundantly regulate flowering in Arabidopsis.Plant Cell 24, 3248-3263. |
| 27 |
de Lucas M, Prat S ( 2014). PIFs get BRright: phytochrome interacting factors as integrators of light and hormonal signals. New Phytol 202, 1126-1141.
DOI URL PMID |
| 28 |
Devlin PF, Kay SA ( 2000). Cryptochromes are required for phytochrome signaling to the circadian clock but not for rhythmicity. Plant Cell 12, 2499-2510.
DOI URL |
| 116 |
Yeom M, Kim H, Lim J, Shin AY, Hong S, Kim JI, Nam HG ( 2014). How do phytochromes transmit the light quality information to the circadian clock in Arabidopsis? Mol Plant 7, 1701-1704.
DOI URL PMID |
| 117 |
Yu JW, Rubio V, Lee NY, Bai SL, Lee SY, Kim SS, Liu LJ, Zhang YY, Irigoyen ML, Sullivan JA, Zhang Y, Lee I, Xie Q, Paek NC, Deng XW ( 2008). COP1 and ELF3 control circadian function and photoperiodic flowering by regula- ting GI stability. Mol Cell 32, 617-630.
DOI URL PMID |
| 29 |
Dotto M, Gómez MS, Soto MS, Casati P ( 2018). UV-B radiation delays flowering time through changes in the PRC2 complex activity and miR156 levels in Arabidopsis tha-liana.Plant Cell Environ 41, 1394-1406.
DOI URL PMID |
| 30 |
Endo M, Tanigawa Y, Murakami T, Araki T, Nagatani A ( 2013). PHYTOCHROME-DEPENDENT LATE-FLOWE- RING accelerates flowering through physical interactions with phytochrome B and CONSTANS. Proc Natl Acad Sci USA 110, 18017-18022.
DOI URL |
| 118 |
Yuan S, Zhang ZW, Zheng C, Zhao ZY, Wang Y, Feng LY, Niu GQ, Wang CQ, Wang JH, Feng H, Xu F, Bao F, Hu Y, Cao Y, Ma LG, Wang HY, Kong DD, Xiao W, Lin HH, He YK ( 2016 a). Arabidopsis cryptochrome 1 functions in nitrogen regulation of flowering. Proc Natl Acad Sci USA 113, 7661-7666.
DOI URL PMID |
| 119 |
Yuan WY, Luo X, Li ZC, Yang WN, Wang YZ, Liu R, Du JM, He YH ( 2016b). A cis cold memory element and a trans epigenome reader mediate Polycomb silencing of FLC by vernalization in Arabidopsis.Nat Genet 48, 1527-1534.
DOI URL PMID |
| 31 | Favory JJ, Stec A, Gruber H, Rizzini L, Oravecz A, Funk M, Albert A, Cloix C, Jenkins GI, Oakeley EJ, Seidlitz HK, Nagy F, Ulm R ( 2009). Interaction of COP1 and UVR8 regulates UV-B-induced photomorphogenesis and stress acclimation in Arabidopsis. EMBO J 28, 591-601. |
| 32 |
Fornara F, Panigrahi KCS, Gissot L, Sauerbrunn N, Rühl M, Jarillo JA, Coupland G ( 2009). Arabidopsis DOF trans- cription factors act redundantly to reduce CONSTANS expression and are essential for a photoperiodic flowering response.Dev Cell 17, 75-86.
DOI URL PMID |
| 33 | Fujiwara S, Oda A, Yoshida R, Niinuma K, Miyata K, Tomozoe Y, Tajima T, Nakagawa M, Hayashi K, Coupland G, Mizoguchi T ( 2008). Circadian clock proteins LHY and CCA1 regulate SVP protein accumulation to control flowe- ring in Arabidopsis. Plant Cell 20, 2960-2971. |
| 34 | Galvão VC, Horrer D, Küttner F, Schmid M ( 2012). Spatial control of flowering by DELLA proteins in Arabidopsis thaliana.Development 139, 4072-4082. |
| 35 |
Gruber H, Heijde M, Heller W, Albert A, Seidlitz HK, Ulm R ( 2010). Negative feedback regulation of UV-B-induced photomorphogenesis and stress acclimation in Arabidopsis. Proc Natl Acad Sci USA 107, 20132-20137.
DOI URL PMID |
| 36 | Guo HW, Yang HY, Mockler TC, Lin CT ( 1998). Regulation of flowering time by Arabidopsis photoreceptors. Science 279, 1360-1363. |
| 120 |
Zuo ZC, Liu HT, Liu B, Liu XM, Lin CT ( 2011). Blue light-dependent interaction of CRY2 with SPA1 regulates COP1 activity and floral initiation in Arabidopsis. Curr Biol 21, 841-847.
DOI URL PMID |
| [1] | 雷雅婷, 王爽, 房伟民, 陈素梅, 陈发棣, 蒋甲福. 切花菊金扇芽变成花受阻的转录组分析[J]. 植物学报, 2026, 61(2): 250-263. |
| [2] | 牛钰凡, 陈玲玲, 苏晨, 王雷. 成花素FT:环境感知与发育调控的信号枢纽[J]. 植物学报, 2026, 61(1): 1-8. |
| [3] | 吴玉俊, 李英菊, 罗巧玉, 马永贵. 光控植物免疫: 从光信号通路到免疫应答的调控网络[J]. 植物学报, 2025, 60(5): 786-803. |
| [4] | 周鑫宇, 刘会良, 高贝, 卢妤婷, 陶玲庆, 文晓虎, 张岚, 张元明. 新疆特有濒危植物雪白睡莲繁殖生物学研究[J]. 植物生态学报, 2025, 49(10): 1643-1655. |
| [5] | 景艳军, 林荣呈. 蓝光受体CRY2化身“暗黑舞者”[J]. 植物学报, 2024, 59(6): 878-882. |
| [6] | 罗燕, 刘奇源, 吕元兵, 吴越, 田耀宇, 安田, 李振华. 拟南芥光敏色素突变体种子萌发的光温敏感性[J]. 植物学报, 2024, 59(5): 752-762. |
| [7] | 杨继轩, 王雪霏, 顾红雅. 西藏野生拟南芥开花时间变异的遗传基础[J]. 植物学报, 2024, 59(3): 373-382. |
| [8] | 陈艳晓, 李亚萍, 周晋军, 解丽霞, 彭永彬, 孙伟, 和亚男, 蒋聪慧, 王增兰, 郑崇珂, 谢先芝. 拟南芥光敏色素B氨基酸位点突变对其结构与功能的影响[J]. 植物学报, 2024, 59(3): 481-494. |
| [9] | 顾家琦, 朱福慧, 谢沛豪, 孟庆营, 郑颖, 张献龙, 袁道军. 棉属光敏色素PHY基因家族的全基因组鉴定与驯化选择分析[J]. 植物学报, 2024, 59(1): 34-53. |
| [10] | 许亚楠, 闫家榕, 孙鑫, 王晓梅, 刘玉凤, 孙周平, 齐明芳, 李天来, 王峰. 红光和远红光在调控植物生长发育及应答非生物胁迫中的作用[J]. 植物学报, 2023, 58(4): 622-637. |
| [11] | 李聪, 齐立娟, 谷晓峰, 李继刚. 植物光信号途径重要新调控因子TZP的研究进展[J]. 植物学报, 2022, 57(5): 579-587. |
| [12] | 杨小凤, 李小蒙, 廖万金. 植物开花时间的遗传调控通路研究进展[J]. 生物多样性, 2021, 29(6): 825-842. |
| [13] | 张长生, 魏滔, 周玉萍, 范甜, 吕天晓, 田长恩. FLC调控植物成花的分子机制研究新进展[J]. 植物学报, 2021, 56(6): 651-663. |
| [14] | 张亚洲, 王淞伟, 何小芳, 杨扬, 陈建国, 孙航. 高山垫状植物团状福禄草开花面积与方位随海拔的变化及其适应性[J]. 植物生态学报, 2020, 44(11): 1154-1163. |
| [15] | 胡文昭, 赵骥民, 张彦文. 二态混合交配系统的适合度优势及其维持机制研究进展[J]. 生物多样性, 2019, 27(4): 468-474. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
 首页
首页