
植物学报 ›› 2022, Vol. 57 ›› Issue (2): 182-196.DOI: 10.11983/CBB21189 cstr: 32102.14.CBB21189
徐海霞, 何静, 易航, 王丽( )
)
收稿日期:2021-11-08
接受日期:2022-02-25
出版日期:2022-03-01
发布日期:2022-03-24
通讯作者:
王丽
作者简介:*E-mail: yzxj@vip.163.com基金资助:
Haixia Xu, Jing He, Hang Yi, Li Wang()
Received:2021-11-08
Accepted:2022-02-25
Online:2022-03-01
Published:2022-03-24
Contact:
Li Wang
摘要: 地钱(Marchantia polymorpha)作为雌雄异株的苔纲植物, 具有对重金属胁迫敏感的特性, 且种群中雌雄比例差异较大, 是研究重金属环境下植物性二态的理想对象之一。为探究雌雄地钱转录组对重金属镉的响应机制差异, 利用高通量测序和加权基因共表达网络分析(WGCNA), 鉴定了在镉胁迫条件下与不同性别地钱有关的模块和基因。结果表明, 镉胁迫响应转录因子雌雄差异主要集中于bHLH、ERF和MYB家族。关键差异响应基因包括MARPO_0147s0038、MARPO_1326s0001和MARPO_0015s0058。差异响应通路雌性主要集中在信号转导通路, 雄性则集中于类黄酮生物合成。研究结果有助于揭示雌雄异株地钱对镉胁迫的反应过程和响应机制, 可为理解雌雄异株植物对重金属胁迫的性别特异性响应机制提供参考。
徐海霞, 何静, 易航, 王丽. 镉胁迫下地钱转录组的性别特异性响应机制. 植物学报, 2022, 57(2): 182-196.
Haixia Xu, Jing He, Hang Yi, Li Wang. Sex Specific Response Mechanism of Transcriptome in Both Male and Female Marchantia polymorpha Under Cadmium Stress. Chinese Bulletin of Botany, 2022, 57(2): 182-196.
| Gene names | Primer sequences (5'-3') |
|---|---|
| MpPCS | F: GAGTCCTTTCGAGCGAATTCT |
| R: AATCTTCCAGCAGTAGCAGTAG | |
| MpActin | F: AGGCATCTGGTATCCACGAG |
| R: ACATGGTCGTTCCTCCAGAC | |
| MARPO_0086s0070 | F: ATTGTCACTTCAAACTGCTTCC R: CTTTGCCAGTATGCTTTCAACT |
| MARPO_0265s0002 | F: TGGCGACTCTTAAAATAGCTCT R: GAAAGTATGGGATTCTTGCGAC |
| MARPO_0001s0061 | F: GAGGAAAATCAACGATACAGGC R: CTTCTCTTGCAGTCAGTTGTTC |
| MARPO_0145s0031 | F: CGTAGGGAGATCAGTGATTACC R: CCTTGTATAACTCACGATCGGA |
| MARPO_0003s0122 | F: TGGACAGAAAACTCATTGGTCT R: ACTTCCTTCTCTCTCACCAATG |
| MARPO_0003s0268 | F: GACTAGGGTTCCTTGGTTCTAC R: CATAATCGGGAAAACGTAACCC |
| MARPO_0011s0046 | F: CGGCTCAGATGTTGATAAATCA R: ATGCACTTCGCTGATCTCATAA |
| MARPO_0032s0085 | F: CCCCTCATCTCAAACTACTTGT R: TTGGAAAAGGAATTGCTGGATG |
表1 qRT-PCR引物序列
Table 1 Primer sequences for qRT-PCR
| Gene names | Primer sequences (5'-3') |
|---|---|
| MpPCS | F: GAGTCCTTTCGAGCGAATTCT |
| R: AATCTTCCAGCAGTAGCAGTAG | |
| MpActin | F: AGGCATCTGGTATCCACGAG |
| R: ACATGGTCGTTCCTCCAGAC | |
| MARPO_0086s0070 | F: ATTGTCACTTCAAACTGCTTCC R: CTTTGCCAGTATGCTTTCAACT |
| MARPO_0265s0002 | F: TGGCGACTCTTAAAATAGCTCT R: GAAAGTATGGGATTCTTGCGAC |
| MARPO_0001s0061 | F: GAGGAAAATCAACGATACAGGC R: CTTCTCTTGCAGTCAGTTGTTC |
| MARPO_0145s0031 | F: CGTAGGGAGATCAGTGATTACC R: CCTTGTATAACTCACGATCGGA |
| MARPO_0003s0122 | F: TGGACAGAAAACTCATTGGTCT R: ACTTCCTTCTCTCTCACCAATG |
| MARPO_0003s0268 | F: GACTAGGGTTCCTTGGTTCTAC R: CATAATCGGGAAAACGTAACCC |
| MARPO_0011s0046 | F: CGGCTCAGATGTTGATAAATCA R: ATGCACTTCGCTGATCTCATAA |
| MARPO_0032s0085 | F: CCCCTCATCTCAAACTACTTGT R: TTGGAAAAGGAATTGCTGGATG |

图1 镉处理后雌性(A)和雄性(B)地钱形态 设置处理后0、24、48、96和168小时5个时间节点。Bars=2 cm
Figure 1 Morphology of female (A) and male (B) Marchantia polymorpha after cadmium (Cd) treatment Five time points after Cd treatment: 0, 24, 48, 96 and 168 h. Bars=2 cm
| Sample_ID | Raw reads | Clean reads | Error (%) | Q20 (%) | Q30 (%) | GC (%) | Total mapped (%) |
|---|---|---|---|---|---|---|---|
| CK_F1 | 50740902 | 50191272 | 0.0241 | 98.38 | 95.02 | 50.92 | 94.74 |
| CK_F2 | 58049938 | 57506790 | 0.0242 | 98.36 | 94.95 | 50.98 | 89.75 |
| CK_F3 | 53602814 | 53058498 | 0.0245 | 98.23 | 94.63 | 51.23 | 95.59 |
| CK_M1 | 49826850 | 49313610 | 0.0243 | 98.31 | 94.84 | 51.14 | 94.22 |
| CK_M2 | 52605578 | 51996578 | 0.0245 | 98.25 | 94.67 | 51.13 | 91.95 |
| CK_M3 | 50064978 | 49523028 | 0.0242 | 98.36 | 94.96 | 50.94 | 90.51 |
| Cd_F1 | 53604084 | 53084914 | 0.0241 | 98.40 | 95.03 | 51.02 | 94.65 |
| Cd_F2 | 48343374 | 47896356 | 0.0255 | 97.87 | 93.60 | 50.53 | 95.16 |
| Cd_F3 | 48556392 | 48058072 | 0.0247 | 98.15 | 94.43 | 50.90 | 96.26 |
| Cd_M1 | 51066022 | 50567610 | 0.0244 | 98.28 | 94.75 | 50.66 | 87.88 |
| Cd_M2 | 51079094 | 50542790 | 0.0244 | 98.28 | 94.75 | 50.48 | 90.25 |
| Cd_M3 | 54825492 | 54241196 | 0.0249 | 98.06 | 94.19 | 50.61 | 93.78 |
表2 转录组质控数据
Table 2 Transcriptome quality control data
| Sample_ID | Raw reads | Clean reads | Error (%) | Q20 (%) | Q30 (%) | GC (%) | Total mapped (%) |
|---|---|---|---|---|---|---|---|
| CK_F1 | 50740902 | 50191272 | 0.0241 | 98.38 | 95.02 | 50.92 | 94.74 |
| CK_F2 | 58049938 | 57506790 | 0.0242 | 98.36 | 94.95 | 50.98 | 89.75 |
| CK_F3 | 53602814 | 53058498 | 0.0245 | 98.23 | 94.63 | 51.23 | 95.59 |
| CK_M1 | 49826850 | 49313610 | 0.0243 | 98.31 | 94.84 | 51.14 | 94.22 |
| CK_M2 | 52605578 | 51996578 | 0.0245 | 98.25 | 94.67 | 51.13 | 91.95 |
| CK_M3 | 50064978 | 49523028 | 0.0242 | 98.36 | 94.96 | 50.94 | 90.51 |
| Cd_F1 | 53604084 | 53084914 | 0.0241 | 98.40 | 95.03 | 51.02 | 94.65 |
| Cd_F2 | 48343374 | 47896356 | 0.0255 | 97.87 | 93.60 | 50.53 | 95.16 |
| Cd_F3 | 48556392 | 48058072 | 0.0247 | 98.15 | 94.43 | 50.90 | 96.26 |
| Cd_M1 | 51066022 | 50567610 | 0.0244 | 98.28 | 94.75 | 50.66 | 87.88 |
| Cd_M2 | 51079094 | 50542790 | 0.0244 | 98.28 | 94.75 | 50.48 | 90.25 |
| Cd_M3 | 54825492 | 54241196 | 0.0249 | 98.06 | 94.19 | 50.61 | 93.78 |
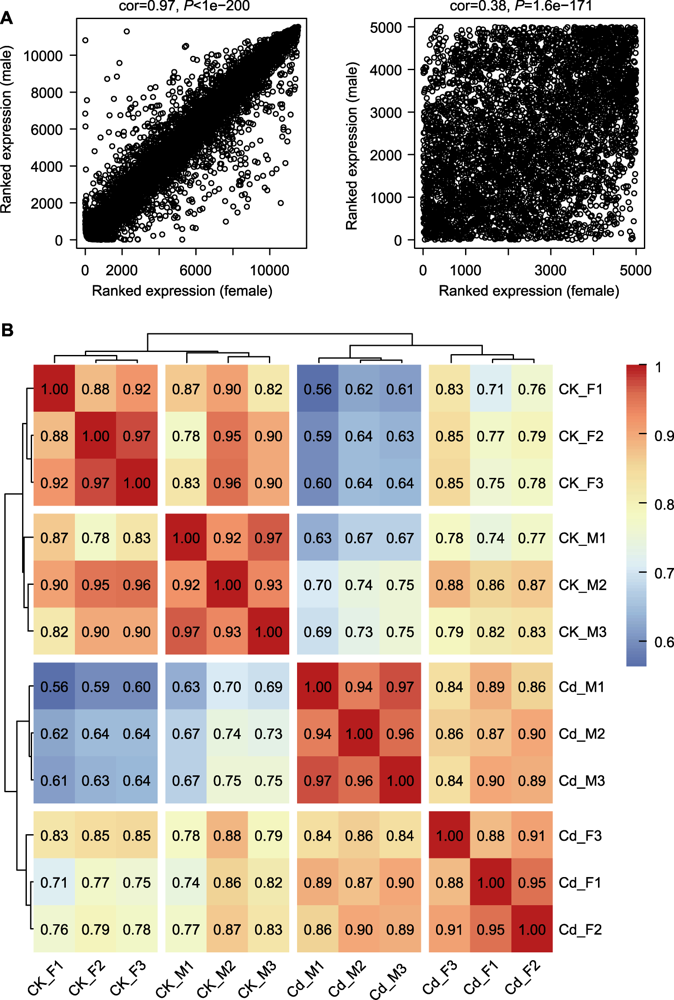
图2 雌雄地钱样本间相关性分析 (A) 雌性和雄性样本间平均基因表达水平与总体连接度的相关性散点图(雌性作为x轴, 雄性作为y轴); (B) 样本间相关性的聚类热图
Figure 2 Correlation analysis among female and male Marchantia polymorpha samples (A) Scatter plot of correlation about average level of gene expression and overall connectivity between female and male samples (females as x-axis and males as y-axis); (B) Clustered heatmap of correlation among samples
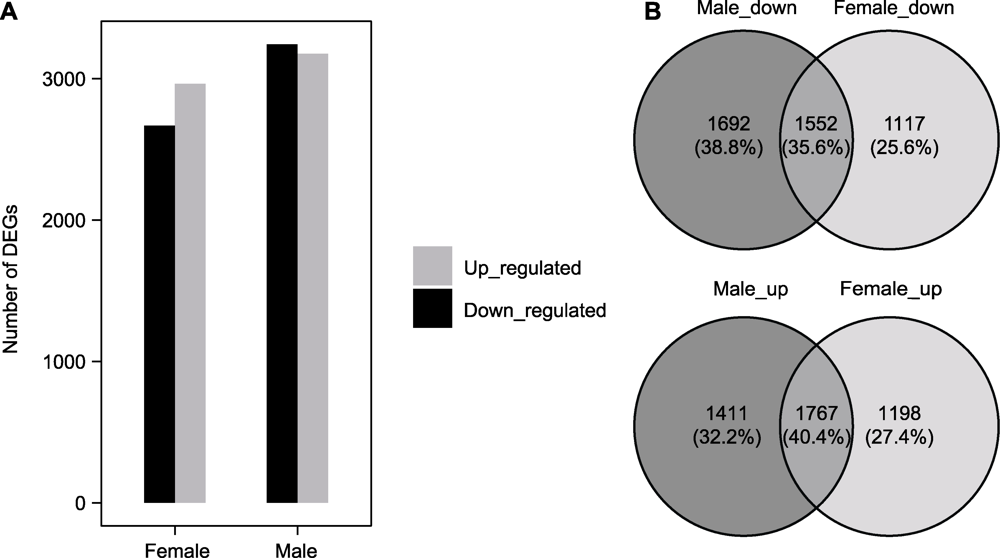
图3 雌雄地钱响应镉胁迫差异表达基因统计 (A) 差异表达基因(DEGs)数目柱状图; (B) 不同变化趋势的差异基因数目韦恩图
Figure 3 Statistics of differentially expressed genes between female and male Marchantia polymorpha in response to cadmium stress (A) Histograms of differentially expressed genes (DEGs); (B) Venn diagram of number of differential genes with different changing trends
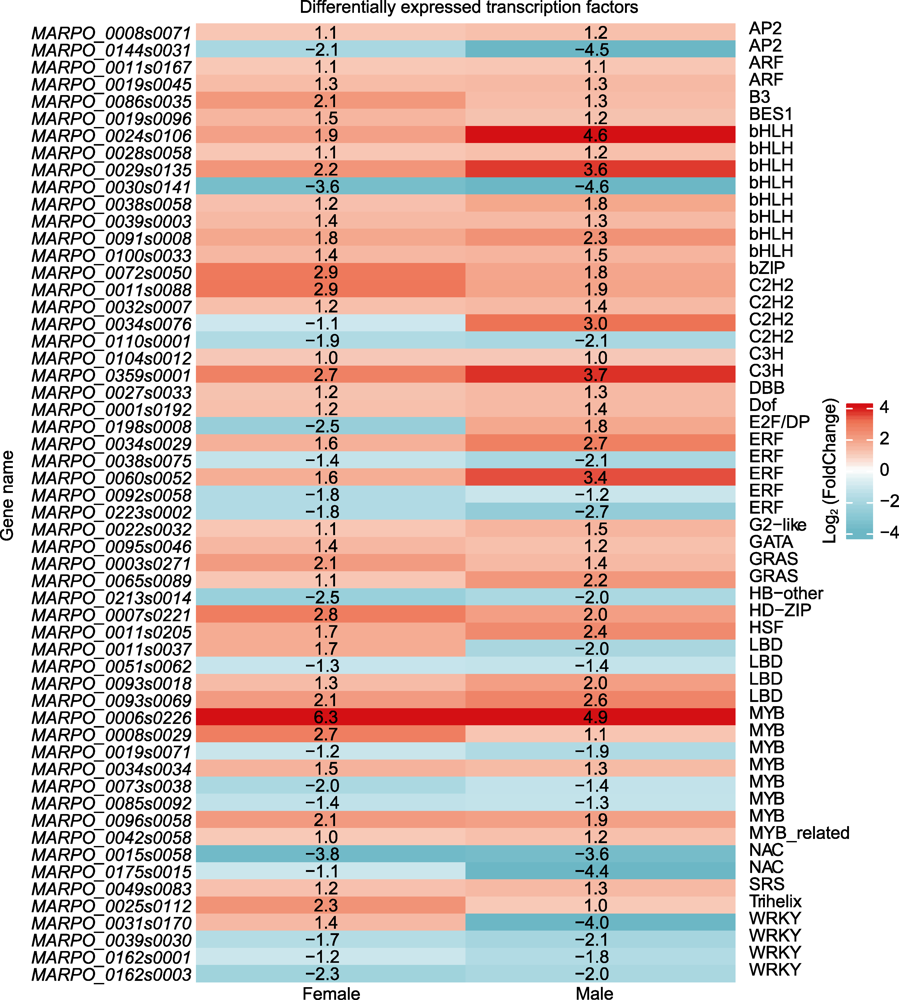
图4 雌雄地钱响应镉胁迫差异表达转录因子
Figure 4 Differentially expressed transcription factors between female and male Marchantia polymorpha in response to cadmium stress
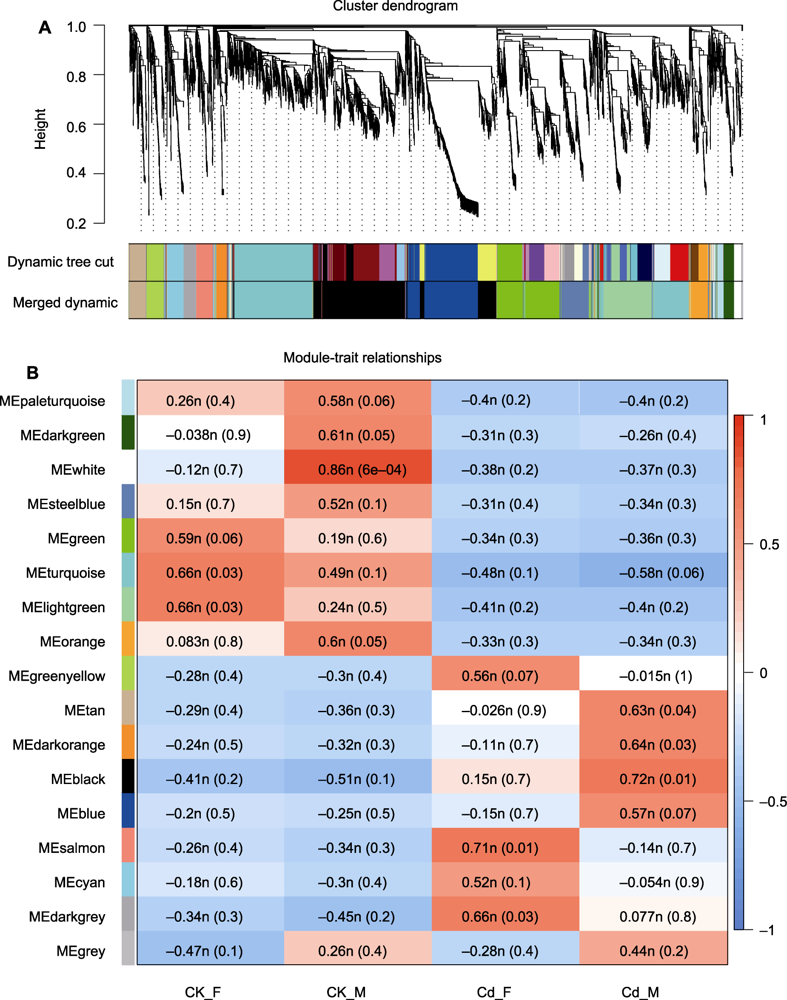
图5 雌雄地钱响应镉胁迫的加权基因共表达网络分析 (A) 基因与模块的聚类树状图; (B) 模块与表型之间的相关性热图。红色表示正相关, 蓝色表示负相关。
Figure 5 Weighted gene co-expression network analysis of female and male Marchantia polymorpha response to cadmium stress (A) Cluster dendrogram of genes and modules; (B) Heatmap of correlation between modules and phenotypes. Red indicates positive correlation and blue indicates negative correlation.
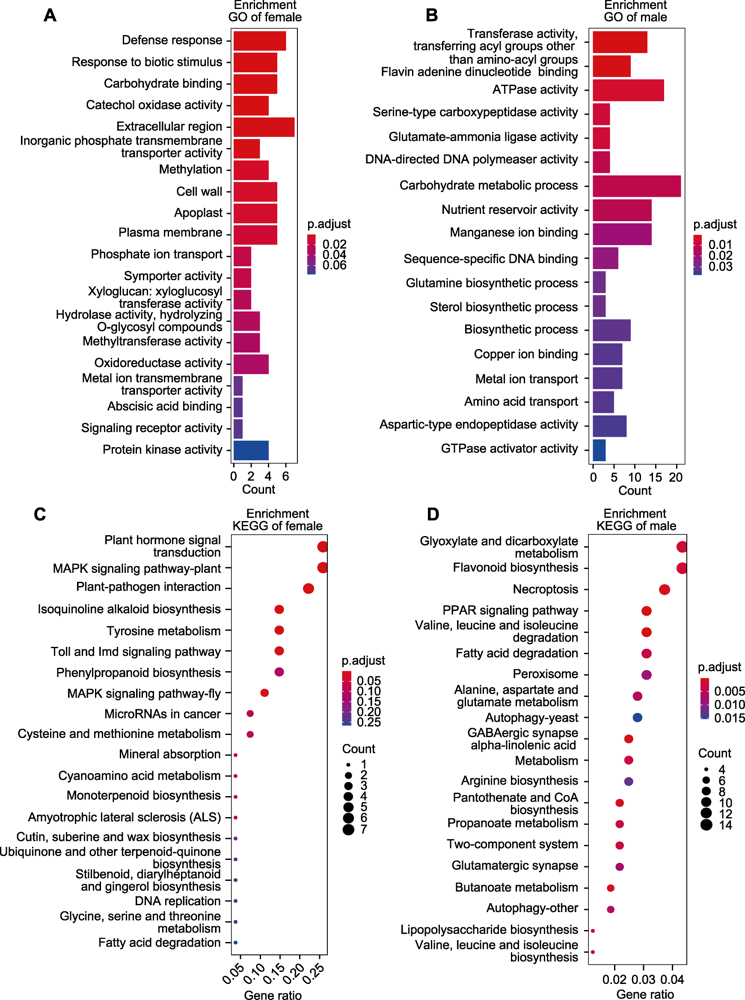
图6 目标模块的富集分析 (A) salmon模块前10条GO条目富集结果; (B) black模块前10条GO条目富集结果; (C) salmon模块前10条KEGG条目富集结果; (D) black模块前10条KEGG条目富集结果
Figure 6 Enrichment analysis of target modules (A) Top ten GO enrichment of salmon module; (B) Top ten GO enrichment of black module; (C) Top ten KEGG enrichment of salmon module; (D) Top ten KEGG enrichment of black module
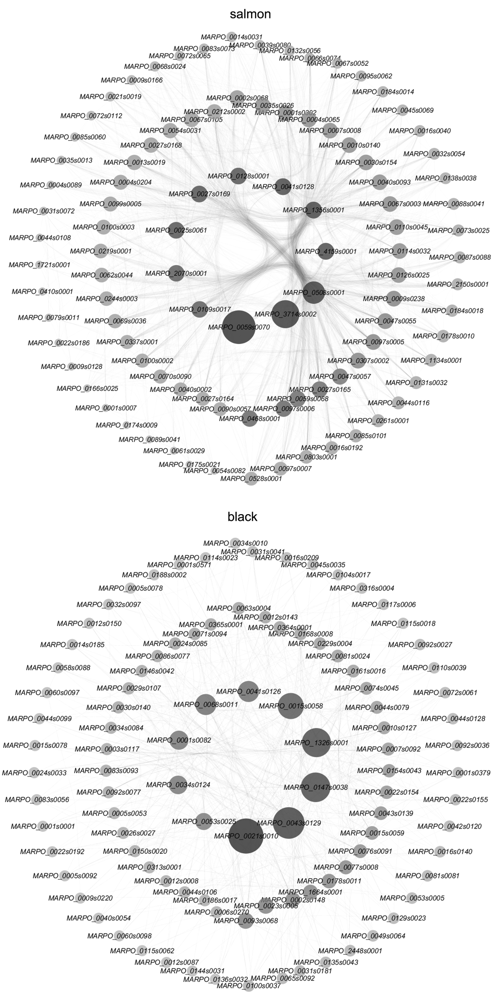
图7 salmon模块和black模块的共表达网络图和核心基因 圆点直径从小到大、从浅色到深色代表degree逐渐增大。
Figure 7 Co-expression network diagram and hub genes of the salmon module and black module The diameter of dot from small to large, with light to dark colors represents the increasement in degree.
| Module | Gene names | Protein names |
|---|---|---|
| black | MARPO_0147s0038 | Phytocyanin domain-containing protein |
| black | MARPO_1326s0001 | ANK_REP_REGION domain-containing protein |
| black | MARPO_0015s0058 | NAC domain-containing protein |
| black | MARPO_0068s0011 | Hydrolase_4 domain-containing protein |
| black | MARPO_0034s0124 | Protein kinase domain-containing protein |
| salmon | MARPO_4159s0001 | Protein kinase domain-containing protein (fragment) |
| salmon | MARPO_0097s0006 | SCP domain-containing protein |
| salmon | MARPO_0047s0057 | FAD-binding FR-type domain-containing protein |
| salmon | MARPO_0009s0238 | S-protein homolog |
| salmon | MARPO_0047s0055 | FAD-binding FR-type domain-containing protein |
表3 salmon模块和black模块中核心基因功能注释
Table 3 Functional annotation of hub genes in salmon module and black module
| Module | Gene names | Protein names |
|---|---|---|
| black | MARPO_0147s0038 | Phytocyanin domain-containing protein |
| black | MARPO_1326s0001 | ANK_REP_REGION domain-containing protein |
| black | MARPO_0015s0058 | NAC domain-containing protein |
| black | MARPO_0068s0011 | Hydrolase_4 domain-containing protein |
| black | MARPO_0034s0124 | Protein kinase domain-containing protein |
| salmon | MARPO_4159s0001 | Protein kinase domain-containing protein (fragment) |
| salmon | MARPO_0097s0006 | SCP domain-containing protein |
| salmon | MARPO_0047s0057 | FAD-binding FR-type domain-containing protein |
| salmon | MARPO_0009s0238 | S-protein homolog |
| salmon | MARPO_0047s0055 | FAD-binding FR-type domain-containing protein |
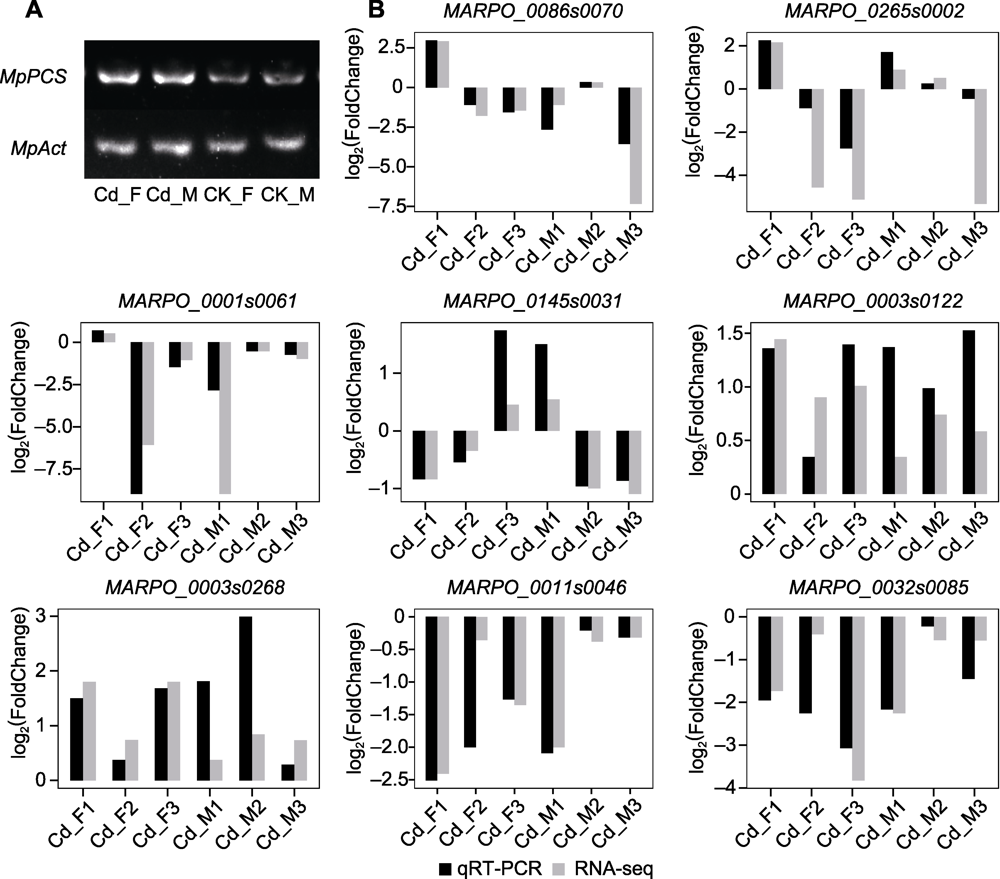
图8 基因表达半定量分析及表达模式验证 (A) 半定量RT-PCR检测4个样本中marker基因(MpPCS)的转录, MpPCS为33个PCR周期, MpAct (作为内参基因)为27个PCR周期; (B) 通过qRT-PCR验证表达数据
Figure 8 Semi-quantitative analysis of gene expression and expression pattern validation (A) Semi-quantitative RT-PCR was used to detect transcription of marker genes (MpPCS) in four samples, 33 PCR cycles for MpPCS, 27 PCR cycles for MpAct (as an internal reference gene); (B) The expression data were verified by qRT-PCR
| [1] | 安婷婷, 黄帝, 王浩, 张一, 陈应龙 (2021). 植物响应镉胁迫的生理生化机制研究进展. 植物学报 56, 347-362. |
| [2] | 王研, 贾博为, 孙明哲, 孙晓丽 (2021). 野生大豆耐逆分子调控机制研究进展. 植物学报 56, 104-115. |
| [3] |
Arai H, Yanagiura K, Toyama Y, Morohashi K (2019). Genome-wide analysis of MpBHLH12, a IIIf basic helix-loop-helix transcription factor of Marchantia polymorpha. J Plant Res 132, 197-209.
DOI URL |
| [4] |
Ares Á, Itouga M, Kato Y, Sakakibara H (2018). Differential metal tolerance and accumulation patterns of Cd, Cu, Pb and Zn in the liverwort Marchantia polymorpha L. Bull Environ Contam Toxicol 100, 444-450.
DOI URL |
| [5] |
Baruah S, Bora MS, Sharma P, Deb P, Sarma KP (2017). Understanding of the distribution, translocation, bioaccumulation, and ultrastructural changes of Monochoria hastata plant exposed to cadmium. Water Air Soil Pollution 228, 17.
DOI URL |
| [6] |
Bisang I, Hedenäs L (2005). Sex ratio patterns in dioicous bryophytes re-visited. J Bryol 27, 207-219.
DOI URL |
| [7] |
Brown RC, Lemmon BE, Shimamura M, Villarreal JC, Renzaglia KS (2015). Spores of relictual bryophytes: diverse adaptations to life on land. Rev Palaeobot Palynol 216, 1-17.
DOI URL |
| [8] |
Degola F, De Benedictis M, Petraglia A, Massimi A, Fattorini L, Sorbo S, Basile A, di Toppi LS(2014). A Cd/Fe/Zn-responsive phytochelatin synthase is constitutively present in the ancient liverwort Lunularia cruciata (L.) Dumort. Plant Cell Physiol 55, 1884-1891.
DOI URL |
| [9] | Elzanati O, Mouzeyar S, Roche J (2020). Dynamics of the transcriptome response to heat in the moss, Physcomitrella patens. Int J Mol Sci 21, 1512. |
| [10] | Fan CH, Gao Yl, Du B (2016). Response of FTIR and Raman spectra on cell wall of calendula of ficinalis seedlings roots to the co-contamination stress of lead and cadmium in loess. Spectrosc Spect Anal 36, 2076-2081. |
| [11] | Jiao Y, Ge W, Qin R, Sun BL, Jiang WS, Liu DH (2012). Influence of cadmium stress on growth, ultrastructure and antioxidative enzymes in Populus 2025. Fresenius Environ Bull 21, 1375-1384. |
| [12] | Kazemitabar SK, Faraji S, Najafi-Zarrini H (2020). Identification and in silico evaluation of bHLH genes in the Sesamum indicum genome: growth regulation and stress dealing specially through the metal ions homeostasis and flavonoid biosynthesis. Gene Rep 19, 100639. |
| [13] |
Li J, Lu HL, Liu JC, Hong HL, Yan CL (2015). The influence of flavonoid amendment on the absorption of cadmium in Avicennia marina roots. Ecotoxicol Environ Saf 120, 1-6.
DOI URL |
| [14] |
Li JY, Zhang XC, Li D, Sun MY, Shi L (2020a). Energy response patterns to light spectrum at sex differentiation stages of Drynaria roosii gametophytes. Environ Exp Bot 172, 103996.
DOI URL |
| [15] |
Li MG, Barbaro E, Bellini E, Saba A, di Toppi LS, Varotto C (2020b). Ancestral function of the phytochelatin synthase C-terminal domain in inhibition of heavy metal-mediated enzyme overactivation. J Exp Bot 71, 6655-6669.
DOI URL |
| [16] |
Ligrone R, Duckett JG, Renzaglia KS (2012). Major transitions in the evolution of early land plants: a bryological perspective. Ann Bot 109, 851-871.
DOI URL |
| [17] | Liu YK, Liu LX, Qi JH, Dang PY, Xia TS (2019). Cadmium activates ZmMPK3-1 and ZmMPK6-1 via induction of reactive oxygen species in maize roots. Biochem Biophys Res Commun 516, 747-752. |
| [18] |
Lo JC, Tsednee M, Lo YC, Yang SC, Hu JM, Ishizaki K, Kohchi T, Lee DC, Yeh KC (2016). Evolutionary analysis of iron (Fe) acquisition system in Marchantia polymorpha. New Phytol 211, 569-583.
DOI URL |
| [19] |
Markert B, Weckert V (1993). Time-and-site integrated long-term biomonitoring of chemical elements by means of mosses. Toxicol Environ Chem 40, 43-56.
DOI URL |
| [20] |
Marks RA, Smith JJ, Cronk Q, Grassa CJ, McLetchie DN (2019). Genome of the tropical plant Marchantia inflexa: implications for sex chromosome evolution and dehydration tolerance. Sci Rep 9, 8722.
DOI PMID |
| [21] |
Muhammad T, Zhang J, Ma YL, Li YS, Zhang F, Zhang Y, Liang Y (2019). Overexpression of a mitogen-activated protein kinase SlMAPK3 positively regulates tomato tolerance to cadmium and drought stress. Molecules 24, 556.
DOI URL |
| [22] |
Paasch AE, Mishler BD, Nosratinia S, Stark LR, Fisher KM (2015). Decoupling of sexual reproduction and genetic diversity in the female-biased mojave desert moss Syntrichia caninervis (Pottiaceae). Int J Plant Sci 176, 751-761.
DOI URL |
| [23] |
Piper PW, Truman AW, Millson SH, Nuttall J (2006). Hsp90 chaperone control over transcriptional regulation by the yeast Slt2(Mpk1)p and human ERK5 mitogen-activated protein kinases (MAPKs). Biochem Soc Trans 34, 783-785.
DOI URL |
| [24] |
Renner SS, Ricklefs RE (1995). Dioecy and its correlates in the flowering plants. Am J Bot 82, 596-606.
DOI URL |
| [25] | Rio DC, Ares M Jr, Hannon GJ, Nilsen TW (2010). Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb Protoc 2010, pdb.prot5439. |
| [26] |
Rottenberg A (1998). Sex ratio and gender stability in the dioecious plants of Israel. Bot J Linn Soc 128, 137-148.
DOI URL |
| [27] |
Rubinstein CV, Gerrienne P, de la Puente GS, Astini RA, Steemans P (2010). Early Middle Ordovician evidence for land plants in Argentina (eastern Gondwana). New Phytol 188, 365-369.
DOI PMID |
| [28] |
Siedlecka A, BaszyńAski T (1993). Inhibition of electron flow around photosystem I in chloroplasts of Cd-treated maize plants is due to Cd-induced iron deficiency. Physiol Plant 87, 199-202.
DOI URL |
| [29] |
Sierocka I, Alaba S, Jarmolowski A, Karlowski WM, Szweykowska-Kulinska Z (2020). The identification of differentially expressed genes in male and female gametophytes of simple thalloid liverwort Pellia endiviifolia sp. B using an RNA-seq approach. Planta 252, 21.
DOI PMID |
| [30] |
Sigfridsson KGV, Bernát G, Mamedov F, Styring S (2004). Molecular interference of Cd2+ with photosystem II. Biochim Biophys Acta 1659, 19-31.
PMID |
| [31] |
Thamm A, Saunders TE, Dolan L (2020). MpFEW RHIZOIDS 1 miRNA-mediated lateral inhibition controls rhizoid cell patterning in Marchantia polymorpha. Curr Biol 30, 1905-1915.
DOI PMID |
| [32] |
Wan LC, Zhang HY (2012). Cadmium toxicity: effects on cytoskeleton, vesicular trafficking and cell wall construction. Plant Signal Behav 7, 345-348.
DOI URL |
| [33] |
Wang JX, Zhang XQ, Shi ML, Gao LJ, Niu XF, Te RG, Chen L, Zhang WW (2014). Metabolomic analysis of the salt-sensitive mutants reveals changes in amino acid and fatty acid composition important to long-term salt stress in Synechocystis sp. PCC 6803. Funct Integr Genomics 14, 431-440.
DOI URL |
| [34] |
Wang Q, Zeng XN, Song QL, Sun Y, Feng YJ, Lai YC (2020). Identification of key genes and modules in response to cadmium stress in different rice varieties and stem nodes by weighted gene co-expression network analysis. Sci Rep 10, 9525.
DOI PMID |
| [35] |
Wu MX, Luo Q, Zhao Y, Long Y, Liu SL, Pan YZ (2018). Physiological and biochemical mechanisms preventing Cd toxicity in the new hyperaccumulator Abelmoschus manihot. J Plant Growth Regul 37, 709-718.
DOI URL |
| [36] |
Xin XY, Chen WH, Wang B, Zhu F, Li Y, Yang HL, Li JG, Ren DT (2018). Arabidopsis MKK10-MPK6 mediates red- light-regulated opening of seedling cotyledons through phosphorylation of PIF3. J Exp Bot 69, 423-439.
DOI URL |
| [37] |
Zhang R, Liu JY, Liu QS, He HG, Xu X, Dong TF (2019). Sexual differences in growth and defence of Populus yunnanensis under drought stress. Can J Forest Res 49, 491-499.
DOI |
| [38] | Zhang ZQ, Hu XN, Zhang YQ, Miao ZY, Xie C, Meng XZ, Deng J, Wen JQ, Mysore KS, Frugier F, Wang T, Dong JL (2016). Opposing control by transcription factors MYB61 and MYB3 increases freezing tolerance by relieving C-repeat binding factor suppression. Plant Physiol 172, 1306-1323. |
| [39] |
Zvereva EL, Kozlov MV (2011). Impacts of industrial polluters on bryophytes: a meta-analysis of observational studies. Water Air Soil Pollut 218, 573-586.
DOI URL |
| [1] | 柯嘉雯, 程张浩, 高雪夷, 徐云剑, 王毅. 镉污染下的植物响应:从吸收、转运到应答与缓解机制[J]. 植物生态学报, 2026, 50(预发表): 1-. |
| [2] | 仝淼, 王欢, 张文双, 王超, 宋建潇. 重金属污染土壤中细菌抗生素抗性基因分布特征[J]. 生物多样性, 2025, 33(3): 24101-. |
| [3] | 张向歌, 陈晨, 程珊, 李春鑫, 朱雅婧, 许欣然, 王会伟. 油莎豆块茎特异性表达基因鉴定及分析[J]. 植物学报, 2025, 60(1): 33-48. |
| [4] | 王亚萍, 包文泉, 白玉娥. 单细胞转录组学在植物生长发育及胁迫响应中的应用进展[J]. 植物学报, 2025, 60(1): 101-113. |
| [5] | 张雅琦, 戎福喜, 沈雨欣, 洪哲源, 张蓝天, 武亮. 植物HIPP家族蛋白结构和功能研究进展[J]. 植物学报, 2024, 59(4): 659-670. |
| [6] | 蔡淑钰, 刘建新, 王国夫, 吴丽元, 宋江平. 褪黑素促进镉胁迫下番茄种子萌发的调控机理[J]. 植物学报, 2023, 58(5): 720-732. |
| [7] | 白雪, 李玉靖, 景秀清, 赵晓东, 畅莎莎, 荆韬羽, 刘晋汝, 赵鹏宇. 谷子及其根际土壤微生物群落对铬胁迫的响应机制[J]. 植物生态学报, 2023, 47(3): 418-433. |
| [8] | 刘厶瑶, 李柱, 柯欣, 孙丽娜, 吴龙华, 赵杰杰. 贵州省典型汞铊矿区周边农田土壤跳虫群落特征[J]. 生物多样性, 2022, 30(12): 22265-. |
| [9] | 张雅楠, 黄蕾, 李佳彬, 张雷, 党振华. 基于转录组的阿拉善地区多枝柽柳多态Genic-SSRs的识别与开发[J]. 植物学报, 2021, 56(4): 433-442. |
| [10] | 刘润, 张朝晖, 申家琛, 王智慧. 岩溶洞穴苔藓群落特征及其对重金属污染的指示意义: 以贵州织金洞为例[J]. 生物多样性, 2018, 26(12): 1277-1288. |
| [11] | 陈权, 马克明. 互花米草入侵对红树林湿地沉积物重金属累积的效应与潜在机制[J]. 植物生态学报, 2017, 41(4): 409-417. |
| [12] | 刘勉, 张彩飞, 黄建勋, 马红. 利用低拷贝核基因重建菊科紫菀亚科族间系统发育关系[J]. 植物学报, 2015, 50(5): 549-564. |
| [13] | 伍自力, 余孟瑶, 陈露, 魏静, 王晓琴, 胡勇, 闫妍, 万平. 小立碗藓对重金属镉胁迫的应答特征[J]. 植物学报, 2015, 50(2): 171-179. |
| [14] | 李琦, 籍霞, 王恩辉, 高寒梅, 衣艳君. 苔藓植物对青岛市大气重金属污染的生物监测作用[J]. 植物学报, 2014, 49(5): 569-577. |
| [15] | 魏志琴, 陈志勇, 秦蓉, 王宇涛, 李韶山. Cu2+对拟南芥根的局部毒性及诱导DNA损伤和细胞死亡[J]. 植物学报, 2013, 48(3): 303-312. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
 首页
首页