
|
|
||
|
重要林木樟科植物全基因组测序研究进展
植物学报
2024, 59 (2):
302-318.
DOI: 10.11983/CBB23035
近年来, 随着测序技术的革新、测序成本的降低和生物信息学软件的开发, 植物全基因组研究蓬勃发展。樟科(Lauraceae)隶属被子植物木兰类, 泛热带分布, 物种多样性高, 其中很多物种具有重要的经济和生态价值, 目前已发表包括8个物种的13个基因组。该文从樟科全基因组研究现状、基因组特征、起源和进化以及功能基因和基因家族4个方面进行综述, 着重介绍基于组学数据的木兰类及樟科的系统发生、樟科经历的多倍化事件以及与樟科花器官进化和代谢产物相关的基因鉴定。结合研究现状展望了樟科基因组研究的发展方向, 建议通过增加测序基因组分支的代表性并关注具有特殊价值的物种, 及研究物种特异性功能基因以加深对该家族基因功能和进化的理解。 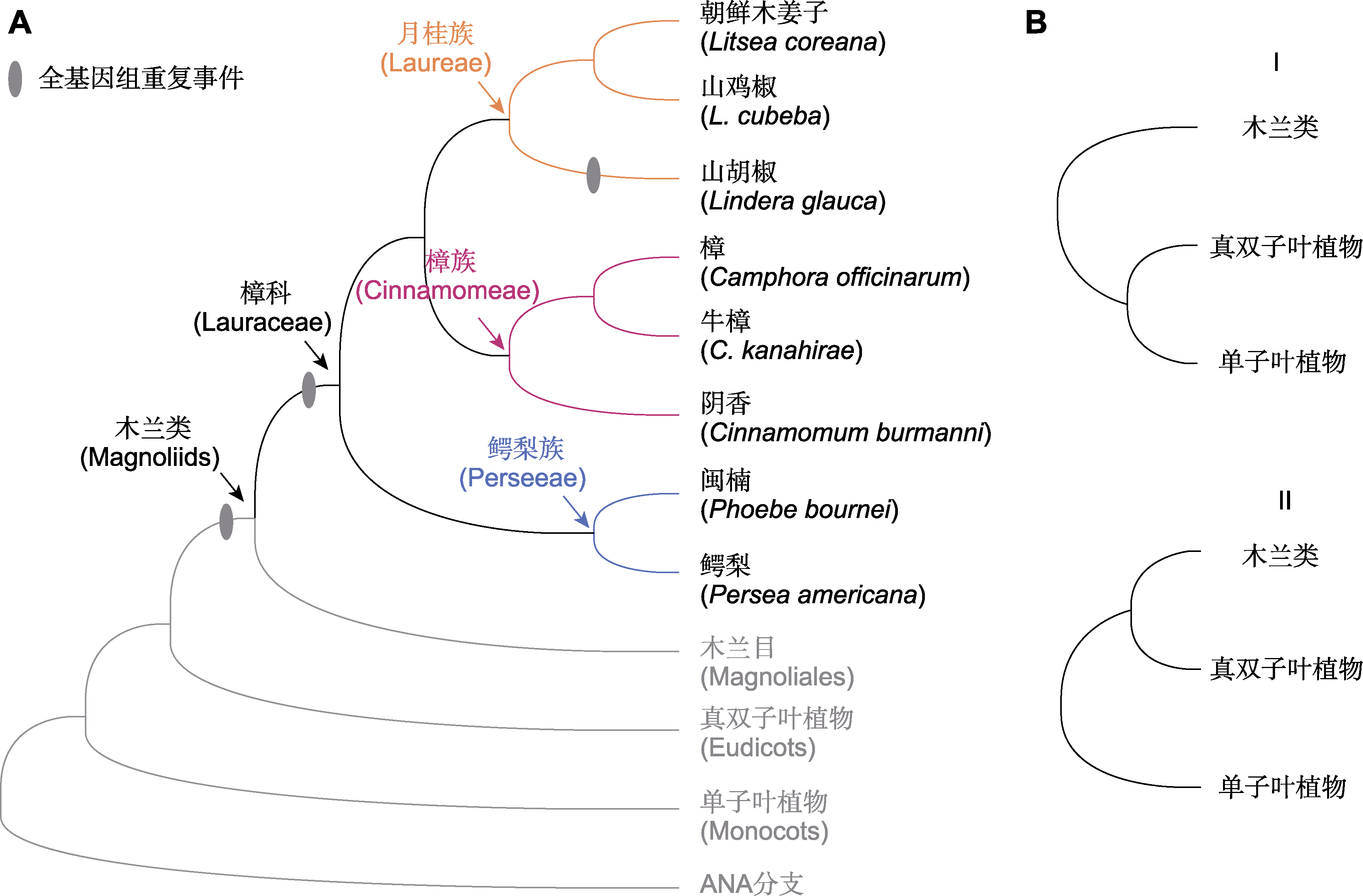 View image in article
图5
樟科已测序物种系统发生关系
(A) 樟科已测序物种系统发生树(参考Liu et al.,
正文中引用本图/表的段落
通过基因组测序获取大量的单拷贝核基因数据并构建生命之树是探讨类群之间系统发生关系的重要方式(王伟和刘阳, 2020)。木兰类、真双子叶植物和单子叶植物之间的进化关系一直困扰着植物学家(Guo et al., 2023)。尽管已有许多木兰类基因组发表, 也利用转录组、叶绿体和线粒体数据构建了被子植物系统发生树(One Thousand Plant Transcriptomes Initiative, 2019; Li et al., 2021; Hu et al., 2023a), 但木兰类、真双子叶植物和单子叶植物之间的进化关系仍然存在争议(Shen et al., 2022)。樟科作为木兰类最大的科(The Angiosperm Phylogeny Group, 2016), 木兰类的系统发生位置也是樟科基因组研究的重点内容之一。樟科基因组相关文章中有11篇对该问题进行了讨论(占84.6%)。在樟科基因组研究结果中, 木兰类的系统发生位置呈现2种结构(图5B): I类是真双子叶植物的姐妹群, II类是真双子叶植物和单子叶植物构成分支的姐妹群。其中结构I更常见, 得到7篇文章的支持。
(2) 增加测序基因组分支的代表性。目前樟科测序的基因组仅涉及3个族, 还有6个族尚无代表物种的基因组发表(图2; 表1)。樟科具有复杂的进化历史, 从系统发生拓扑结构不一致中可窥探一二。有代表性的物种取样能够帮助厘清樟科的系统发生关系。注释良好的基因组数据为使用重测序和靶向捕获的方法从标本中获取大量的基因序列提供参考, 这将解决樟科采集和鉴定难题, 从而构建更具有代表性的系统发生树(Sun et al., 2022a)。一方面, 解析樟科系统发生关系不仅可帮助我们从进化角度探讨特定功能基因的生物学特性和分子机制, 还能为保护濒危植物提供理论依据。桫椤(Alsophila spinulosa)基因组就是1个良好的案例(Huang et al., 2022)。研究者基于9个群体107个桫椤个体的基因组重测序解析树蕨类植物树干形成发育的独特性, 为理解蕨类植物的进化提供了新视角, 也从影响桫椤种群变化的因素中拓展了保护工作思路。另一方面, 基于系统发生树和分布数据可揭示樟科的时空分布格局, 解析其类群多样分布类型的成因。例如, 泛热带分布的厚壳桂属和琼楠属(Beilschmiedia) (Chanderbali et al., 2001), 东亚-北美间断分布的檫木属(Sassafras) (Yang et al., 2022b; Qin et al., 2023), 亚洲-大洋洲分布的土楠属(Endiandra) (Li et al., 2020), 以及仅分布在亚洲的新樟属(Neocinnamomun)和仅分布在美洲的桂土楠群(Chanderbali et al., 2001; Liu et al., 2021)。此外, 大量遗传信息和代表性取样能够为解析樟科复杂的进化历史提供支持(Liu et al., 2021; Tian et al., 2021)。目前, 樟科科内的多倍化只在山胡椒基因组中检测到(图5), 考虑到月桂族多样的染色体变化及其更大的基因组(图3), 增加月桂族物种全基因组测序, 有助于理解多倍化在樟科进化中的作用。
本文的其它图/表
|
 首页
首页


{kind=link}