
|
|
||
|
P515的测量原理、方法和应用
植物学报
2021, 56 (5):
594-604.
DOI: 10.11983/CBB21052
光谱技术已广泛应用于光合研究领域, 如光吸收信号P515和P700氧化还原动力学以及叶绿素荧光等, 可快速、准确地检测植物的光合活性。P515信号广泛存在于高等植物和藻类中, 是类囊体膜上的色素分子吸收光能后, 其吸收光谱发生位移造成。利用光诱导的P515快速和慢速动力学, 可检测PSI和PSII反应中心的比值、ATP合酶的质子传导性、围绕PSI的环式电子传递速率、质子动力势及其组分, 还可通过同步检测叶绿素荧光和P515信号研究光保护机制。该文总结了P515的主要测量原理、方法及其应用, 旨在为深入研究光合作用机理提供技术支持。 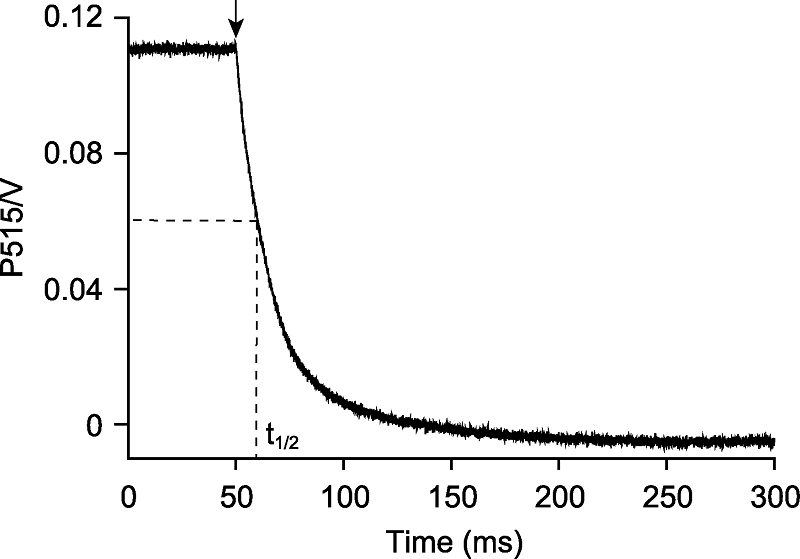 View image in article
图2
测量拟南芥叶绿体ATP合酶的质子传导性
拟南芥离体叶片在光强为500 μmol·m-2·s-1的活化光(635 nm)下照光10分钟, 照光期间提供稳定的CO2 (380 μmol·mol-1)、湿度(60%大气湿度)和温度(23°C)。检测黑暗250 ms的P515衰减动力学, 重复100次后取平均值。ATP合酶的质子传导率为P515衰减动力学曲线半衰期(t1/2)的倒数, 即94.3·s-1。箭头表示活化光关闭。
正文中引用本图/表的段落
当植物处于稳态的光合作用时, 通过光合电子传递引起的质子流入类囊体囊腔的速率与通过ATP合酶的质子流出速率相等(Sacksteder and Kramer, 2000)。DIRK (dark internal relaxation kinetics)技术是利用短暂的黑暗(<500 ms)干扰处于稳态光合的电子和质子流, 检测样品在520 nm处的光吸收(Sacksteder et al., 2000)。该方法可估算叶绿体ATP合酶的活性(Cruz et al., 2001)。其原理是当ATP合酶活性降低时, 通过ATP合酶的质子流出速率也会相应降低。拟南芥野生型在500 μmol·m-2·s-1的光照强度下, 叶绿体ATP合酶的质子传导率(gH+)为94.3·s-1 (图2)。拟南芥bfa1和bfa3是两个ATP合酶突变体, 在不同光照强度下, 其gH+分别下降至野生型的30%和50%, 同时均表现出较高的NPQ (Zhang et al., 2016, 2018)。这是因为当ATP合酶活性降低时, 通过ATP合酶的质子流出效率降低, 导致质子在类囊体囊腔累积, 囊腔过度酸化会抑制NPQ衰减, 从而表现出高水平的NPQ。烟草(Nicotiana tabacum) GTG-atpB突变体的gH+降至野生型的25%, 且突变体的碳同化能力、线式电子传递速率和P700+再还原能力也显著降低(Rott et al., 2011)。P515与叶绿素荧光、P700氧化还原动力学的结合使用在烟草、玉米(Zea mays)、拟南芥和豌豆(Pisum sativum)等叶绿体ATP合酶突变体的表型筛选和功能研究方面发挥了重要作用(Rott et al., 2011; Zoschke et al., 2012; Fristedt et al., 2015; Sukhov et al., 2016; Zhang et al., 2016)。
本文的其它图/表
|
 首页
首页
{kind=link}