|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
一个新的黄瓜叶色突变体鉴定、初定位及转录组分析
植物学报
2025, 60 (4):
515-532.
DOI: 10.11983/CBB24112
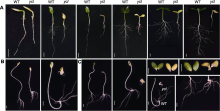
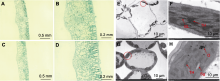
叶色突变体是研究光形态发生、叶绿体发育、叶绿素代谢和光合作用机制等多种生理过程的理想材料。该研究从黄瓜(Cucumis sativus) XYYH-2-1-1株系自交后代中获得1个新的黄化致死突变体ycl (yellow cotyledon lethal)。该突变体自幼苗出土后子叶一直呈黄化状态, 约2周后枯萎死亡, 其生长抑制表型为非光依赖型。与野生型相比, ycl突变体的Chl a和Chl b含量趋于零, 叶绿素生物合成途径中Mg2+螯合过程受阻。显微和超微结构分析发现, ycl叶片组织紊乱、叶绿体发育受阻。ycl的抗氧化酶活性及丙二醛含量显著升高, 说明其受到氧化胁迫, 且抗氧化能力强。ycl净光合速率极显著降低, 胞间CO2浓度上升, 推测ycl光合速率降低源于气孔导度降低、叶绿素含量减少和叶绿体发育受阻。转录组学分析表明, ycl与其野生型间存在337个差异表达基因, 光合作用、类黄酮生物合成、叶绿素代谢和活性氧代谢是导致ycl黄化致死表型形成的关键途径。通过BSA-Seq分析, ycl突变基因初步定位于3号染色体的1.48-1.9 Mb区间, 内含41个候选基因。对ycl突变体的研究为阐明黄瓜叶绿体发育的分子机制提供了参考。
表5
ycl突变位点初定位区间内基因
正文中引用本图/表的段落
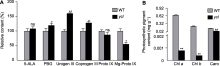
为了进一步探讨突变体中光合色素缺失的原因, 对叶绿素生物合成中间代谢产物含量进行测定。将野生型叶绿素合成的各种前体物质含量统一设定为100%, 结果表明, 与野生型相比, 突变体子叶中5-氨基乙酰丙酸(5-ALA)、胆色素原(PBG)、尿卟啉原III以及粪卟啉原III的含量均上升, 分别为野生型的107.56%、118.99%、158.35%和127.31%; 而原卟啉IX及Mg-原卟啉IX的含量有所降低, 为野生型的86.25%和55.31%, 但Proto IX的含量与野生型相比无显著差异(图2A)。由此推测ycl突变体叶绿素合成过程中Proto IX镁离子螯合受阻, 导致Mg-Proto IX积累减少, 使得合成通路无法合成后续产物, 最终使叶绿素a和叶绿素b含量急剧降低。
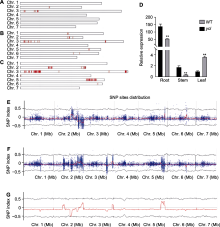
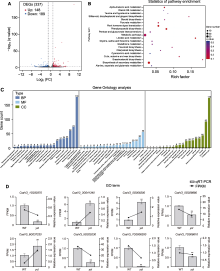
BSA混池测序结果显示, 在第18号群体中获得15 323个差异SNP, 在XY子叶群体中获得18 953个差异SNP, 在XY真叶群体中获得20 973个差异SNP, 在3个群体中获得野生亲本纯合、隐性池差异且纯合、显性池纯合或杂合的位点分别有19、51和730个SNP, 以1 000 000 bp为窗口, 100 000 bp为步长, 根据隐性池减去显性池的SNP index作图(图5), 发现3号染色体上SNP密集, 且在3个群体中有重叠。由于与目标性状关联的SNP在染色体上连锁遗传, 故将目的基因初定位在第3号染色体的1.48-1.9 Mb区间, 此候选区段中包含41个蛋白编码基因(表5), 后续将进一步精细定位以获得候选突变位点。
取ycl及野生型子叶期(7天)植株根、茎和叶, 利用荧光定量PCR对候选区间内CsaV3_3G001980基因进行表达量分析, 发现在根和茎中, ycl基因的表达量极显著低于野生型; 在叶中, 其表达量极显著高于野生型(图5)。CsaV3_3G001980编码NPF (NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER FAMI- LY)蛋白, 与拟南芥NRT1.5编码基因同源, NRT1.5参与硝酸盐从根到茎的木质部运输, 推测ycl突变体中该基因的表达量下降会影响硝酸盐的运输, 导致叶色发生变化。候选基因CsaV3_3G001980序列全长为6 364 bp, 设计包含启动子区在内的CsaV3_3G001980 gDNA引物, 分段进行克隆测序, 对测序数据拼接并比对, 结果证实未发生基因突变。
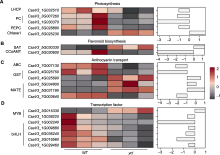
为进一步揭示黄化致死表型形成的原因, 对高度相关的差异表达基因进行筛选分析。在转录组差异表达基因中, 发现5个参与光合作用的关键基因(图7A), 分别为1个编码叶绿素a/b结合蛋白(chlorophyll a/b binding protein, LHCP)的基因、2个编码质体蓝素(plastocyanin, PC)的基因、1个编码磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)且富集于光合生物中的碳固定基因和1个编码叶绿素酶1 (Chlase 1)的基因。与野生型相比, 前4个基因在突变体ycl中表达下调, 后1个基因表达上调。
类黄酮被划分为六大类, 其在植物体内以黄酮、查尔酮、花青素和前花青素等多种形式存在, 类黄酮物质不仅影响植株的呈色, 还是植物抗逆过程中的重要物质。本研究中, 通过转录组测序发现类黄酮生物合成通路上存在2个DEGs (图7B), 其中编码黄酮3′,5′-甲基转移酶(如咖啡酰辅酶A-3-O甲基转移酶 (trans-caffeoyl-CoA 3-O-methyltransferase, CCoAMT))的基因上调, 编码Vinorine合成酶(如司替木丹碱-O-乙酰转移酶(stemmadenine O-acetyltransferase, SAT))的基因显著下调, 说明突变体ycl中黄酮类物质的合成代谢受到调控, 影响了叶片呈色。前人研究发现, 花青素完成聚合需要先与转运蛋白结合。多药和有毒物排出家族(multidrug and toxic compound extrusion, MATE)转运蛋白和谷胱甘肽S-转移酶(glutathione-S-transferase, GST)被证实参与花青素的转运过程, GST还在植物生长发育和非生物胁迫响应中发挥多种作用。我们在转录组差异表达基因中发现5个参与此过程的关键基因, 2个基因编码GST, 3个基因编码MATE转运蛋白(图7C), 编码ABC转运蛋白的基因表达下调也表明突变体中活性氧高度积累, 说明ycl的花青素跨膜或囊泡转运及抗氧化功能受到一定影响。转录因子也可能是导致突变体产生叶色突变的重要指标之一, 转录组测序分析发现ycl及其野生型之间存在7个编码转录因子的差异表达基因(图7D), 5个编码bHLH转录因子, 且全部下调表达, 2个编码MYB转录因子, 说明突变体的花青素含量发生变化, 极有可能影响了叶片呈色。
花青素、叶绿素和类胡萝卜素在叶片中所占的比例及分布直接影响植物叶片的呈色(Jiao et al., 2020; Gao et al., 2021)。我们在ycl中发现5个DEGs参与花青素的转运, 包括维持植物细胞活性氧平衡的ABC转运蛋白(Awai et al., 2006; Zhu et al., 2019)、通过转运原花青素前体参与花青素合成的MATE转运蛋白(Sun et al., 2022), 及参与植物生长发育、次生代谢、逆境抗性和花青素跨膜运输的GST转运蛋白(Jiang et al., 2019; Ghangal et al., 2020)。本文的研究结果表明, ycl的叶色变化与花青素的转运合成途径受调控相关。同时, RNA-Seq分析结果显示, ycl中有2个编码转录因子MYB的差异表达基因, 5个编码bHLH转录因子的差异表达基因且均下调。MYB在植物次生代谢、逆境应答以及形态建成中发挥重要作用(Qiu et al., 2014; Boase et al., 2015; Zhou et al., 2015)。bHLH作为MBW (MYB-bHLH-WD40)复合物中的转录因子, 通过与MYB协同互作激活结构基因以促进原花青素的合成(Allan et al., 2008; Butelli et al., 2008; Zhou et al., 2015; Xie et al., 2016; Dasgupta et al., 2017)。因此, 编码转录因子的DEGs可能导致突变体的花青素含量发生变化; 同时, 下游光合作用和抗氧化基因无法激活, 使代谢紊乱, 最终导致叶色改变。由此可见, 突变体ycl黄化致死表型是多基因层级调控失效的结果, 包括叶绿素代谢失衡、ROS积累与氧化胁迫及转录因子网络失调等。
本文的其它图/表
|
 首页
首页