|
|
|||||||||||||||||||||
|
一个新的黄瓜叶色突变体鉴定、初定位及转录组分析
植物学报
2025, 60 (4):
515-532.
DOI: 10.11983/CBB24112
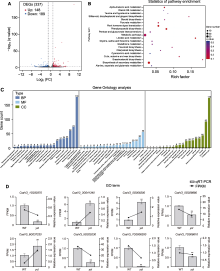
叶色突变体是研究光形态发生、叶绿体发育、叶绿素代谢和光合作用机制等多种生理过程的理想材料。该研究从黄瓜(Cucumis sativus) XYYH-2-1-1株系自交后代中获得1个新的黄化致死突变体ycl (yellow cotyledon lethal)。该突变体自幼苗出土后子叶一直呈黄化状态, 约2周后枯萎死亡, 其生长抑制表型为非光依赖型。与野生型相比, ycl突变体的Chl a和Chl b含量趋于零, 叶绿素生物合成途径中Mg2+螯合过程受阻。显微和超微结构分析发现, ycl叶片组织紊乱、叶绿体发育受阻。ycl的抗氧化酶活性及丙二醛含量显著升高, 说明其受到氧化胁迫, 且抗氧化能力强。ycl净光合速率极显著降低, 胞间CO2浓度上升, 推测ycl光合速率降低源于气孔导度降低、叶绿素含量减少和叶绿体发育受阻。转录组学分析表明, ycl与其野生型间存在337个差异表达基因, 光合作用、类黄酮生物合成、叶绿素代谢和活性氧代谢是导致ycl黄化致死表型形成的关键途径。通过BSA-Seq分析, ycl突变基因初步定位于3号染色体的1.48-1.9 Mb区间, 内含41个候选基因。对ycl突变体的研究为阐明黄瓜叶绿体发育的分子机制提供了参考。
表3
ycl和XYYH-3-1叶片的光合色素含量
正文中引用本图/表的段落
摘要:
叶色突变体是研究光形态发生、叶绿体发育、叶绿素代谢和光合作用机制等多种生理过程的理想材料。该研究从黄瓜(Cucumis sativus) XYYH-2-1-1株系自交后代中获得1个新的黄化致死突变体ycl (yellow cotyledon lethal)。该突变体自幼苗出土后子叶一直呈黄化状态, 约2周后枯萎死亡, 其生长抑制表型为非光依赖型。与野生型相比, ycl突变体的Chl a和Chl b含量趋于零, 叶绿素生物合成途径中Mg2+螯合过程受阻。显微和超微结构分析发现, ycl叶片组织紊乱、叶绿体发育受阻。ycl的抗氧化酶活性及丙二醛含量显著升高, 说明其受到氧化胁迫, 且抗氧化能力强。ycl净光合速率极显著降低, 胞间CO2浓度上升, 推测ycl光合速率降低源于气孔导度降低、叶绿素含量减少和叶绿体发育受阻。转录组学分析表明, ycl与其野生型间存在337个差异表达基因, 光合作用、类黄酮生物合成、叶绿素代谢和活性氧代谢是导致ycl黄化致死表型形成的关键途径。通过BSA-Seq分析, ycl突变基因初步定位于3号染色体的1.48-1.9 Mb区间, 内含41个候选基因。对ycl突变体的研究为阐明黄瓜叶绿体发育的分子机制提供了参考。
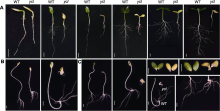
黄瓜(Cucumis sativus L.)黄化致死突变体ycl是通过黄瓜XYYH-2-1-1株系自交性状分离获得, 为自然突变体, 其突变性状稳定遗传。由于突变体在子叶期黄化致死, 突变基因仅能通过杂合体保存, 因此将杂合株系自交产生的后代播种后, 即可分离获得黄化苗, 黄化苗株系可确定为杂合株; XYYH-2-1-1后代中稳定遗传的正常绿叶表型XYYH-3-1则为野生型对照。播种野生型和杂合株系共92株, 构建野生型基因池、杂合体基因池和突变体基因池, 单株取样, 用于突变基因初步定位。以上材料均种植于湖南农业大学实验温室和植物激素与生长发育湖南省重点实验室植物培养室。亲本XYYH-2-1-1由湖南省蔬菜研究所陈惠明研究员惠赠, 于子叶期(播种后7天)对XYYH-3-1及ycl株系的株高、茎粗及叶面积进行评价, 重复3次, 每次重复测量5株。
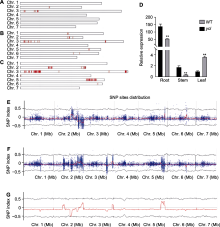
采用混合分组分析(bulked segregation analysis, BSA) (Michelmore et al., 1991)对突变基因进行初定位。采用CTAB法提取基因组DNA并检测质量, 对第18号分离群体进行单基因BSA分析, 取子叶构建2个DNA池: 15棵绿色植株子叶构成18GP基因池、15棵黄化植株子叶构成18YP基因池。对18个群体的所有单株开展BSA-Seq分析, 取子叶构建3个DNA池: 10棵野生型植株子叶构成XYY-WT基因池、8个株系共25棵黄化植株子叶构成XYYpool基因池、8个株系共25棵杂合株系绿色植株子叶构成XYGpool基因池; 因取子叶构建的混池测序获得的差异SNP较少, 而当ycl及其野生型进入真叶期后, 光合作用等相关代谢基因表达水平差异更明显, 故取真叶构建了3个基因池: 10棵野生型植株真叶构成XYY-WTZ基因池、8个株系共25棵黄化植株真叶构成HHZSZ基因池、8个株系共25棵杂合株系绿色植株真叶构成XYGZ基因池。待检验合格后委托湖南拓唯生物科技有限公司进行全基因组重测序, 先使用Qubit2.0软件进行初定量, 随后将文库稀释至1 ng·μL-1并对其insert size进行检测, 用Q-PCR法准确定量文库的有效浓度后使用illumina Novaseq6000仪测序, 得到全基因组测序结果。使用BWA软件将reads比对到黄瓜参考基因组, 使用SAMtools软件寻找全基因组的SNP位点, 最终确定目标基因所在的染色体和大致位置。
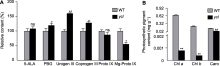
与野生型(XYYH-3-1)相比, 突变体ycl的Chl a、Chl b、Caro和Total Chl含量均显著降低, 分别下降至野生型的0.12%、0.19%、0.14%和20.32%, 并且叶绿素a和叶绿素b的含量均趋于零。ycl叶绿素a/b为1.93, XYYH-3-1叶绿素a/b为3.01, 说明相较于叶绿素b, 突变体的叶绿素a含量下降幅度更大; 此外, 突变体与野生型的总叶绿素/类胡萝卜素(Total Chl/Caro)值分别为0.04和6.63 (图2B; 表3), 表明ycl叶片黄化的形成主要是叶绿素含量显著降低, 尤其是叶绿素a含量减少所致。
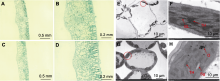
分别对ycl和XYYH-3-1子叶期叶片进行显微观察, 发现野生型叶片中, 海绵组织与栅栏组织结构饱满、排列有序且紧密(图3A, B); 突变体ycl中, 栅栏组织和海绵组织细胞排列散乱, 缺乏明确的界限, 细胞之间的空隙较大, 叶脉也相对较小(图3C, D), 表明ycl的叶片组织发育异常。用透射电镜(transmission electron microscope, TEM)观察ycl突变体和野生型子叶的超微结构, 发现野生型叶片的叶绿体形状大多为长椭圆形或梭形, 细胞内数目多(图3E); ycl突变体叶片的叶绿体膨胀成近圆形(图3G)。与野生型相比, ycl突变体的叶绿体结构简单且发育不良, 基粒片层稀疏, 叶绿体内积累了大量的嗜锇体(图5H), 表明黄化致死突变体ycl的叶绿体发育存在明显缺陷。
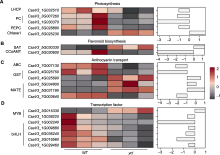
取ycl及野生型子叶期(7天)植株根、茎和叶, 利用荧光定量PCR对候选区间内CsaV3_3G001980基因进行表达量分析, 发现在根和茎中, ycl基因的表达量极显著低于野生型; 在叶中, 其表达量极显著高于野生型(图5)。CsaV3_3G001980编码NPF (NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER FAMI- LY)蛋白, 与拟南芥NRT1.5编码基因同源, NRT1.5参与硝酸盐从根到茎的木质部运输, 推测ycl突变体中该基因的表达量下降会影响硝酸盐的运输, 导致叶色发生变化。候选基因CsaV3_3G001980序列全长为6 364 bp, 设计包含启动子区在内的CsaV3_3G001980 gDNA引物, 分段进行克隆测序, 对测序数据拼接并比对, 结果证实未发生基因突变。
本文的其它图/表
|
 首页
首页