|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
水稻胚乳淀粉合成相关蛋白的翻译后修饰研究进展
植物学报
2025, 60 (2):
256-270.
DOI: 10.11983/CBB24067
蛋白质翻译后修饰(PTMs)是调控蛋白质生物学功能的重要机制, 在水稻(Oryza sativa)种子发育和胚乳淀粉生物合成中起重要作用。随着蛋白质组学的发展, 已在水稻胚乳中鉴定到大量淀粉合成相关酶(SSREs)发生蛋白质PTMs。该文总结了水稻胚乳SSREs的磷酸化、赖氨酸乙酰化、琥珀酰化、2-羟基异丁酰化、丙二酰化及泛素化6种PTMs的蛋白质组学分析、修饰位点和途径及生物学功能。其中, 蛋白质磷酸化修饰的研究最多, 其在调节植物生长发育和淀粉合成代谢过程中起关键作用。此外, 还讨论了PTMs对籽粒灌浆、稻米淀粉品质及外观的潜在作用。该文综述了PTMs在水稻胚乳淀粉合成相关蛋白中的调控机制, 为培育高产优质水稻品种提供了有价值的参考。
表4
水稻种子淀粉合成蛋白赖氨酸乙酰化修饰
正文中引用本图/表的段落
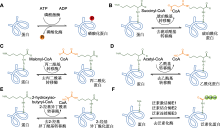
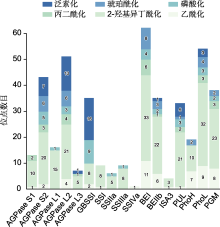
赖氨酸ε-氨基的可逆乙酰化(lysine acetylation, Kac)是一种普遍发生且高度保守的PTMs, 可调节多种细胞生理过程, 包括转录、细胞周期进程和能量代谢(Svensson et al., 2020), 在植物生长发育以及胁迫响应中起重要作用(He et al., 2016; Luo et al., 2017)。赖氨酸乙酰化酶(lysine acetyltransferases)将乙酰基从乙酰辅酶A转移到特定的赖氨酸残基, 形成蛋白质的乙酰化修饰, 而赖氨酸去乙酰化酶(lysine deacetylases)则可以逆转该反应。目前, 已在不同功能和亚细胞定位的拟南芥(Finkemeier et al., 2011)、大豆(Glycine max) (Smith-Hammond et al., 2014)和小麦(Zhang et al., 2016)的蛋白中鉴定出多个Kac修饰位点。功能注释结果表明, 这些乙酰化蛋白除了参与RNA合成和蛋白质加工外, 还参与信号转导以及许多代谢调控过程。水稻胚乳SSREs的AGPase S1、AGPase S2、AGPase L2、GBSSI、SSI、SSIVa、BEI、BEIIb、ISA3、PUL、PhoH、PhoL和磷酸葡萄糖变位酶(phosphoglucomutase, PGM)均被证实会发生Kac, 修饰位点数为1-11个不等(表4)。Wang等(2017)对日本晴未授粉雌蕊及胚乳蛋白进行乙酰化组学分析, 从972个乙酰化蛋白中获得1 688个Kac修饰肽和1 817个Kac修饰位点, 其中包括胚乳淀粉合成以及种子发育相关蛋白, 表明Kac可能在水稻胚乳淀粉生物合成以及种子发育早期具有重要调控作用。Meng等(2018)从水稻种子692个蛋白中鉴定出1 003个Kac位点, 并发现与水稻胚乳淀粉合成途径相关的部分关键酶, 如AGPase S2、AGPase L2、GBSSI、BEI、BEIIb、ISA3、PUL、PhoH及PhoL都发生了较大程度的乙酰化修饰, 推测Kac可能是淀粉代谢所必需。Xue等(2018)从水稻幼苗叶片的866个蛋白质组中检测到1 353个Kac位点, 同时发现乙酰化修饰具有保守性。Xiong等(2016)从水稻716个蛋白中鉴定出位于多种细胞器内的1 337个Kac位点, 这些位点参与调控包括淀粉合成在内的多种生理生化过程以及信号通路。
蛋白质泛素化参与调控高温及干旱胁迫下的水稻发育(郭慧妍等, 2024), 在水稻代谢调节、蛋白质合成与降解等方面发挥关键作用。在高温条件下, Ying等(2023b)利用蛋白质无标记定量技术在2个水稻品种(9311和GLA4)的胚乳中鉴定出246个Kub蛋白以及488个Kub位点。其中与淀粉合成相关的关键酶包括AGPL2、AGPL3、AGPS1、AGPS2、GBSSI、BEI、BEIIb、PUL和Pho1 (表7)。在AGPL2、AGPL3、AGPS1及AGPS2中鉴定出22个Kub位点。高温胁迫下AGPL2多个Kub位点的修饰水平发生变化。在2个水稻品种中AGPL2-K254位点的修饰水平均显著上调, 表明该位点可能在高温下AGPase调控中起关键作用。从GBSSI中鉴定出16个Kub位点, 高温胁迫下2个水稻品种的6个位点(K130、K177、K399、K381、K385和K549)泛素化水平上调, 而9311的1个位点(K258)泛素化水平下调。研究表明, 高温胁迫下水稻直链淀粉含量降低(Pan et al., 2023), 推测可能与GBSSI蛋白泛素化水平显著升高导致其发生降解有关。在BEI中发现3个Kub位点(K103、K108和K122), 高温下9311中K108和K122的泛素化水平显著上调。在BEIIb中仅发现1个Kub位点(K134), 高温下2个水稻品种的泛素化水平均略微升高。推测这些变化可能是高温下胚乳淀粉糊化温度和相对结晶度升高的原因之一。在PUL中发现5个Kub位点(K230、K330、K432、K736和K884), 根据高温胁迫下泛素化水平调控模式与蛋白质丰度调控一致, 推测泛素化修饰位点与蛋白质降解有关, 还可能通过改变酶活性以及蛋白互作来调节支链淀粉的生物合成。在Pho1中发现3个Kub位点(K277、K445和K941), 高温环境下Pho1- K277和Pho1-K445两个位点分别仅在9311和GLA4中被检测到发生泛素化。Pang等(2021)研究表明, 高温胁迫下水稻(特别是GLA4)中Pho1的蛋白丰度降低。因此, 这些仅发生在高温环境中的Kub可能与Pho1蛋白的降解有关。以上研究表明, 在逆境下泛素化修饰对水稻淀粉合成具有重要调控作用。
研究表明植物体内蛋白质PTMs之间存在串扰机制(PTM-crosstalk), 这些多层面的串扰作用在植物生长发育以及胁迫响应等方面具有重要意义。目前, 植物体内不同蛋白质PTMs之间串扰调控机制的研究尚处于起步阶段。庞悦涵等(2024)以水稻胚乳磷酸化和泛素化蛋白质组学数据为基础, 对2种PTMs进行关联分析。结果显示, 在246个泛素化蛋白中有143个同时具有磷酸化修饰位点, 表明在水稻胚乳中这2种PTMs密切相关。进一步分析发现, 在淀粉代谢通路中AGPL2、AGPL3、AGPS1、AGPS2、GBSSI、BEI、BEIIb、PUL及Pho1均具有双重修饰位点, 表明磷酸化与泛素化修饰在水稻胚乳淀粉生物合成过程中有重要调控作用。在高温胁迫下, 9311中Pho1磷酸化水平显著下降而其泛素化水平显著上升。推测泛素化水平升高导致蛋白降解, 由于蛋白表达丰度降低, 磷酸化水平随之下调。GLA4中AGPL2磷酸化修饰和泛素化修饰均具有上调和下调的多个位点, 是唯一同时具有双重修饰和多重调节位点的蛋白。说明多重PTMs在高温胁迫下对淀粉合成具有重要的协同调节作用。
(ac)表示赖氨酸乙酰化位点. ... A comprehensive catalog of the lysine-acetylation targets in rice (Oryza sativa) based on proteomic analyses 1 2016 ... 赖氨酸ε-氨基的可逆乙酰化(lysine acetylation, Kac)是一种普遍发生且高度保守的PTMs, 可调节多种细胞生理过程, 包括转录、细胞周期进程和能量代谢(Svensson et al., Global identification and systematic analysis of lysine malonylation in maize (Zea mays L.) 2 2021 ... 在谷物胚乳生物合成过程中, 多种SSREs通过相互作用形成多酶复合体(multi-enzyme complex, MEC)协同合成淀粉, MEC的组装及活性受蛋白质磷酸化调控(Yu et al.,
本文的其它图/表
|
 首页
首页