|
|
|||||||||||||||||||||||||||||||||||||||||||||||||
|
基因编辑技术在玉米中的研究进展
植物学报
2024, 59 (6):
978-998.
DOI: 10.11983/CBB24080
基因编辑技术已成为现代农业育种领域的重要工具。玉米(Zea mays)是全球最重要的粮食作物之一, 基因编辑技术在玉米中的应用显示出巨大的潜力。该文综述了基因编辑技术在玉米研究中的应用进展, 重点介绍了CRISPR/Cas等系统在玉米基因组编辑中的最新成果。首先, 介绍了基因编辑技术的基本原理和类型, 特别是CRISPR/Cas系统的工作机制及其在玉米中的应用优势。其次, 总结了基因编辑技术在玉米育种中的研究进展, 涵盖从基础的基因组编辑到复杂的多基因编辑, 旨在改良玉米的产量、品质和抗逆性等关键性状。最后, 文章列举了我国在玉米基因编辑方面的杰出工作, 并讨论了基因编辑技术应用于玉米育种中存在的问题, 同时展望了未来发展方向。
表1
CRISPR/Cas及其衍生技术
正文中引用本图/表的段落
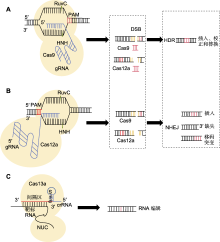
CRISPR/Cas基因编辑技术起源于细菌的免疫系统, 用于抵抗病毒入侵, 通过一系列重复的DNA序列(CRISPR)和相关蛋白质(Cas)进行DNA切割。自2012年在基因编辑中应用以来, 该技术迅速发展成为基因工程领域的核心工具。CRISPR技术有多种类型, 包括CRISPR/Cas9、CRISPR/Cas12a (Cpf1)、CRISPR/ Cas13 (专注于RNA编辑)和CRISPR/Cas的衍生技术BE (base editor)和PE (prime editing)技术。CRISPR/ Cas系统及其衍生技术各具特色与优势(表1), 能够精确且高效地满足多样化的基因编辑需求, 实现对基因组的精准调控与修饰, 并使其在基因治疗、农业改良和疾病模型构建等多个领域展现出巨大的应用潜力。
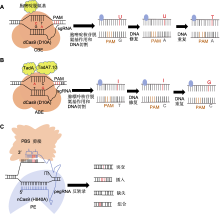
(A) sgRNA识别并结合到目标DNA序列特定的原间隔区邻近基序(PAM)位点, dCas9蛋白则与sgRNA结合, 形成复合物并结合于目标DNA上, 但不切割DNA双链, 仅使其单链化; 随后胞嘧啶脱氨酶在sgRNA的引导下, 接触到暴露的单链DNA上的胞嘧啶, 并催化其脱氨反应, 将胞嘧啶转化为尿嘧啶; 最后在DNA重复或修复过程中, 尿嘧啶被视为胸腺嘧啶的类似物, 从而纳入到新合成的DNA链中, 实现C-G碱基对到T-A碱基对的直接替换; (B) 利用CRISPR-Cas9系统中的sgRNA识别并结合到目标DNA序列的PAM位点, Cas9蛋白与sgRNA结合形成复合物, 并定位于目标DNA上; ABE中的腺嘌呤脱氨酶在sgRNA的引导下, 接触到暴露的单链DNA上的腺嘌呤, 并催化其脱氨反应, 将腺嘌呤转化为次黄嘌呤或脱氧次黄嘌呤; 最后在DNA修复过程中, 识别到次黄嘌呤后, 启动修复过程, 中间产物通常被替换为鸟嘌呤, 从而实现A-T碱基对到G-C碱基对的直接替换; (C) 利用pegRNA作为引导分子结合sgRNA, 并在其3'末端增加引物结合位点(PBS)序列和逆转录模板(RTT); 在pegRNA的引导下, 部分失活的Cas9切口酶切断含PAM序列的DNA单链; 切割后的DNA单链与pegRNA的3'末端PBS序列互补并结合, 随后逆转录酶沿RTT模板序列开始逆转录反应, 将目标编辑序列直接引到DNA切口处; 随后细胞内DNA修复机制识别并处理切口处的DNA结构, 最终保留携带目标编辑的DNA链。CBE、ABE和PE同
目前, 通过基因编辑技术已成功创制了多种玉米核雄性不育和温敏雄性不育突变体。特别是MS45基因, 作为控制花粉发育的关键基因, 其突变导致不育性状能稳定遗传。Svitashev等(2015)通过CRISPR/Cas9对MS45基因进行定点编辑, 获得双等位基因的雄性不育突变体。此外, 通过对MS8基因进行编辑产生雄性不育突变体, 并且这些突变体遵循孟德尔遗传规律(Chen et al., 2018)。研究表明, ZmFAR1和ZmABCG26作为受microRNA调控的脂质代谢关键基因, 在玉米花药发育和脂质代谢中发挥重要作用。通过对ZmABCG26以及脂质代谢相关基因ZmFAR1进行CRISPR/Cas9定点编辑, 在突变体花药中观察到角质层减少而蜡质含量增加的变化, 这有助于创造新的雄性不育系(Jiang et al., 2021)。此外, 通过靶向敲除ZmTMS5基因获得热敏雄性不育玉米品种, 其可在特定的温度下表现出不育性状(Li et al., 2017b)。在植物发育的关键阶段, 小RNA (sRNA)在调控靶向mRNA的转录水平、保持基因组的完整性以及增强DNA甲基化方面发挥重要作用。特别是在许多单子叶植物中, Dcl5 (Dicer-like 5)酶经精确切割mRNA生成24 nt的分阶段次级小干扰RNA (phasiRNAs)。在开花植物的减数分裂过程中, phasiRNAs对花药的正常功能具有关键作用。Teng等(2020)通过CRISPR-Cas9系统生成包括等位基因以及一个转座子中断的等位基因在内的几个dcl5突变体, 并发现dcl5突变体几乎或根本没有24 nt phasiRNAs, 其花药短小且绒毡层细胞有缺陷, 并表现出温敏雄性不育性状。
玉米植株的形态特征如密植耐受性、茎秆强度、植株高度、叶片夹角和穗位高度, 对于玉米光合效率和空间资源利用具有决定性作用, 并直接影响产量。因此, 在玉米育种和农业生产实践中, 株型优化是提高产量和作物适应性的关键因素。在玉米发育初期, 特别是在9至10叶期, 位于叶腋的分生组织(AM)转变为花序分生组织(IM), 继而在IM上发育出多行小穗分生组织(SPM)。而分生组织的生长和发育主要受CLAVATA- WUSCHEL信号途径中CLE肽信号调控, 此途径是果穗行数及粒重增加的关键(Schoof et al., 2000)。Liu等(2021)利用CRISPR/Cas9基因编辑技术, 对玉米中CLV3的同源基因CLE7和FCP1的启动子区进行特定的缺失编辑, 发现突变体中分生组织显著增大, 从而间接提高玉米产量相关性状, 如穗大小、粒数及百粒重。叶夹角是影响玉米密植的关键因子, 优化叶夹角不仅能提高光利用效率, 还可改善作物群体的空气流通, 最终提高群体光合效率。Li等(2017a)利用CRISPR/Cas9对玉米ZmLg1基因进行编辑, 该基因编码一种含SBP (squamosa promoter-binding protein)结构域的蛋白, 负责调控叶夹角。通过对ZmLg1基因第1外显子进行编辑, T0代植株展现出高达51.5%-91.2%的基因编辑效率, 并使叶夹角明显变小至对照组的50%, 且突变特征稳定遗传。田间试验表明, 这些突变体通过增加种植密度展现出增产潜力。SAMBA作为植物特异性调节因子, 是影响拟南芥有丝分裂期推进复合体(APC/C)的关键因子, 负责控制细胞周期的顺利进行。研究发现, SAMBA与APC/C在玉米中同样存在相互作用, 通过对玉米中2个SAMBA基因敲除突变体的观察, 发现一系列生长缺陷, 包括节间缩短、上部叶片缩小和直立以及叶片整体缩小(Gong et al., 2022)。玉米中功能基因brd1和ZmRAVL1通过油菜素内酯信号通路调控叶夹角。UPA2定位于ZmRAVL1上游, 具有与DRL1蛋白结合的能力。LG1蛋白参与调控玉米叶夹角, 能激活ZmRAVL1的表达。DRL1与LG1直接相互作用, 抑制LG1对ZmRAVL1的激活。相较于玉米, 其祖先种大刍草(Zea mays subsp. parviglumis)的UPA2位点与DRL1蛋白结合能力更强, 导致ZmRAVL1表达显著受抑制, 进而下调其下游基因brd1的表达, 降低叶环区内源BR水平, 使叶夹角变小。将大刍草UPA2等位基因导入玉米或对玉米中的ZmRAVL1进行编辑, 在密植条件下均可显著提高玉米的产量(刘杰和严建兵, 2019)。
果穗作为玉米产量形成的关键组分, 其形态特征如穗长、行数及籽粒数直接影响种子的生产潜力。数量性状基因座(quantitative trait locus, QTLs)对穗长起核心控制作用。但目前定位到与穗长相关的QTLs非常少, 且其分子机制大多未知。通过全基因组关联分析, Luo等(2022)鉴定出YIGE1, 其编码一个未知蛋白, 通过影响雌花小花数量调节穗长, 且过表达YIGE1能显著增加雌性花序分生组织的大小, 从而增加穗长和粒数, 提高总体产量。而使用CRISPR/Cas9技术敲除YIGE1或通过高度活跃的DNA转座子(mutator, Mu)插入产生的突变体与之表现相反。穆路遥(2022)基于3个百粒重关键基因Zm079、Zm080和Zm081, 通过CRISPR/Cas9技术成功创制了多份基因编辑材料。Zm080基因敲除导致部分植株百粒重明显降低, 而其它植株的穗重、穗长和穗粗显著增加。对Zm081基因的编辑影响植株的株高、穗位高和雄穗分枝数等。
近年来, 利用基因编辑技术, 特别是CRISPR/Cas9技术提高玉米抗病性已成为研究热点。通过CRISPR/ Cas9技术精确修改与病原体互作的基因, 能够显著提高玉米对特定真菌或病毒的抵抗力。例如, 玉米黑粉菌(Ustilago maydis)是当前最严重的玉米病害之一。Pathi等(2020)发现在黑粉菌侵染后玉米中脂肪氧化酶(lipoxygenase, Lox)基因ZmLox3的表达量显著升高, 表明其编码产物在黑粉菌侵染过程中发挥作用。利用CRISPR/Cas9技术对ZmLox3基因第1外显子进行定点编辑, 得到的突变体植株表现出对黑粉菌的抵抗力增强, 体内黑粉菌生物量显著减少。此外, 玉米粗缩病是由水稻黑条矮缩病毒(rice black- streaked dwarf virus, RBSDV)引起的病害。研究表明, 玉米中ZmGDlα基因编码的RabGDP解离抑制因子在囊泡形成和运输中发挥重要作用, 并被确定为玉米粗缩病的易感因子。Liu等(2022)采用CRISPR/ Cas9技术将编辑位点设定在ZmGDlα基因第1外显子和翻译起始密码子下游30 bp处, 产生1 bp的插入和32 bp的缺失突变体。通过对比接种病毒后的植株表型, 发现野生型植株表现出矮化表型, 叶片背面出现蜡状突起, 且不能正常结实。相反, 基因编辑后的突变体具有良好的生长发育能力和显著抵抗RBSDV的能力。禾谷镰孢菌(Fusarium graminearum)是玉米茎腐病的主要致病菌之一。在植物中, 茉莉酸(jasmonic acid, JA)作为关键的内源激素参与对病害的防御反应。在茉莉酸信号途径中, COI (coronatine-insensitive)蛋白是关键受体, JAZ (jasmonate zim-domain)蛋白则发挥负调控作用。Ma等(2021)利用CRISPR/Cas9技术编辑玉米ZmCOI1a和ZmJAZ15基因, 发现ZmCOI1a-ZmJAZ15作用模块在调控玉米对赤霉穗腐病抗性中起关键作用。在无JAs信号时, ZmJAZ15与转录因子结合, 抑制防御相关基因的表达; 当存在JAs信号时, ZmCOI1a与ZmJAZ15相互作用, 导致ZmJAZ15被泛素化降解, 从而解除对防御相关基因的抑制。由立枯丝核菌(Rhizoctonia solani)引起的纹枯病严重危害玉米生长, 研究人员利用全基因组关联分析(genome wide association study, GWAS)推测ZmFBL41是关键基因。而转座子插入突变体zmfbl41 (mu1059763)较对照表现出更强的抗性, ZmFBL41蛋白是E3泛素连接酶复合体成员, 能介导复合体对肉桂醇脱氢酶的降解, 从而降低木质素的积累, 使玉米易感纹枯病(李伟滔等, 2019)。上述研究成果在抗病育种、病害防控、基础研究和农业可持续发展方面具有重要的应用价值。
本文的其它图/表
|
 首页
首页