
|
|
||
|
玉米雄性不育机理及其在工程核不育制种中的应用
植物学报
2024, 59 (6):
932-949.
DOI: 10.11983/CBB24078
玉米(Zea mays)是我国种植面积最大和总产量最高的第一大粮食作物, 同时也是杂种优势利用的典范。但与发达国家相比, 我国玉米生产仍然存在着平均单产偏低、突破性品种缺乏和杂交种生产成本高等突出问题。雄性不育系的应用可进一步提高玉米杂种优势的利用效率并最终提高单产。该文综述了玉米雄性不育的分类、基因克隆与机理解析以及分子调控网络构建最新研究进展, 系统介绍了已建立的玉米新型工程核不育技术体系及其应用前景, 为推动玉米雄性发育生物学研究与开展玉米雄性不育杂交育种和制种提供重要参考。 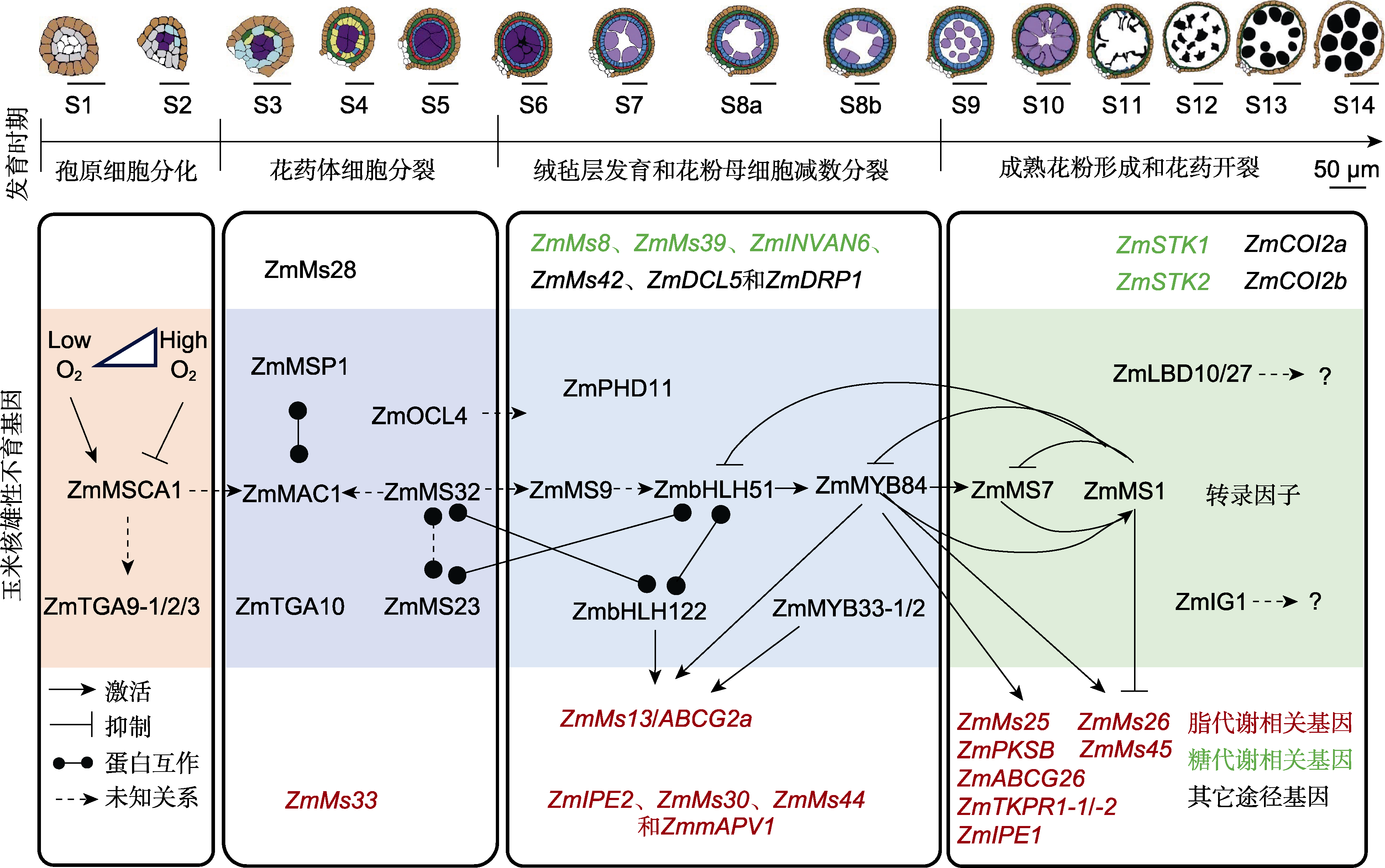 View image in article
图2
已克隆的玉米核不育(GMS)基因及其调控花药和花粉发育的分子网络
正文中引用本图/表的段落
玉米花药和花粉发育进程按照细胞学特征可划分为14个时期(Stages) (S1-S14), 主要包括4个生物学事件: 孢原细胞分化(S1-S2)、花药体细胞分裂(S3- S5)、绒毡层发育和花粉母细胞减数分裂(S6-S8b)以及成熟花粉形成和花药开裂(S9-S14) (Wan et al., 2019)。随着鉴定到玉米GMS基因数目的增加和部分基因分子机理的深入解析, 针对上述4个生物学事件, 已初步搭建了影响玉米花药和花粉发育的分子调控网络(图2)。
花药体细胞经历平周分裂后, 从外到内可形成典型的花药四层壁结构: 表皮层、内皮层、中间层和绒毡层。其中, 包围着发育中小孢子的绒毡层在花粉发育中起重要作用, 它为花粉壁的形成提供了必需的营养和主要成分。绒毡层发育缺陷往往导致花粉畸变和雄性不育(Wan et al., 2019, 2020)。目前, 已鉴定出许多影响玉米花药绒毡层发育和花粉母细胞减数分裂的GMS基因, 针对此生物学事件的转录调控研究也较为详细(图2)。
其次, 5个脂代谢、3个糖代谢和3个其它途径相关的GMS基因在玉米花药发育S6-S8b时期高表达, 同时参与绒毡层发育、花粉母细胞减数分裂、花药角质层和花粉外壁发育(图2)。其中, ZmMs13/ZmABCG2a在花药中特异性表达, 且在S5、S8b和S10期有3个表达峰, 分别受转录因子ZmbHLH122、ZmMYB84和ZmMYB33-1/-2调控, 参与调控胼胝质降解、绒毡层程序性细胞死亡(PCD)、花粉外壁发育以及花药角质层形成(Fang et al., 2023b)。此外, 在该发育阶段高表达的其它GMS基因与上述转录调控通路的关系, 尚待进一步研究。
在玉米中已经鉴定了许多在花药发育后期(S9-S14)表达和发挥功能的GMS基因, 包括5个转录因子、8个脂质代谢相关基因和4个其它途径GMS基因(图2)。其中, PHD-finger类转录因子ZmMS7与玉米核因子(NF-Y)亚基相互作用形成ZmMS7-NF-YA6-YB2-YC9/ 12/15蛋白复合物, 通过直接结合下游靶基因启动子区域的CCAAT盒激活其下游靶基因的表达, 调控花粉外壁和花药角质层的形成(An et al., 2020)。最新研究表明, ZmMs1/ZmLBD30位于ZmbHLH51-ZmMYB84-ZmMS7的转录激活级联下游并被激活表达, 然后作为抑制因子通过反馈抑制关闭该级联激活通路, 通过直接抑制下游脂代谢GMS基因(如ZmMs26和ZmMs45), 或通过抑制ZmMYB84间接抑制下游脂代谢GMS基因(如ZmMs2/ABCG26、ZmTKPR1-1/2、ZmMs25、ZmPKSB、ZmIPE1、ZmMs26和ZmMs45), 以确保花粉发育过程中绒毡层及时退化、合适的孢粉素合成及花粉外壁厚度(Zhang et al., 2021; Liu et al., 2022a; Fang et al., 2023b; Hou et al., 2023)。基因表达和突变体表型分析表明, ZmIG1、ZmLBD10和ZmLBD27参与调控玉米花药发育后期进程, 但其分子机制尚不清楚(Jiang et al., 2021)。
本文的其它图/表
|
 首页
首页
{kind=link}