玉米雄性不育机理及其在工程核不育制种中的应用
吴锁伟, 安学丽, 万向元
植物学报
2024, 59 ( 6):
932-949.
DOI: 10.11983/CBB24078
玉米(Zea mays)是我国种植面积最大和总产量最高的第一大粮食作物, 同时也是杂种优势利用的典范。但与发达国家相比, 我国玉米生产仍然存在着平均单产偏低、突破性品种缺乏和杂交种生产成本高等突出问题。雄性不育系的应用可进一步提高玉米杂种优势的利用效率并最终提高单产。该文综述了玉米雄性不育的分类、基因克隆与机理解析以及分子调控网络构建最新研究进展, 系统介绍了已建立的玉米新型工程核不育技术体系及其应用前景, 为推动玉米雄性发育生物学研究与开展玉米雄性不育杂交育种和制种提供重要参考。
| 序号 | 不育基因 | 基因ID | 编码蛋白 | 参考文献 | | I 转录因子类 | | 1 | ZmMs1/ZmLBD30 | Zm00001d036435 | LBD转录因子 | Hou et al., 2023 | | 2 | ZmIG1 | Zm00001d042560 | LBD转录因子 | Evans, 2007 | | 3 | ZmLBD10 | Zm00001d033335 | LBD转录因子 | Jiang et al., 2021 | | 4 | ZmLBD27 | Zm00001d013732 | LBD转录因子 | Jiang et al., 2021 | | 5 | ZmMs7 | Zm00001d020680 | PHD-finger转录因子 | Zhang et al., 2018a | | 6 | ZmPHD11 | Zm00001d013416 | PHD-finger转录因子 | Jiang et al., 2021 | | 7 | ZmMs9 | Zm00001d028777 | MYB转录因子 | Albertsen et al., 2016 | | 8 | ZmMYB33-1 | Zm00001d012544 | MYB转录因子 | Jiang et al., 2021 | | 9 | ZmMYB33-2 | Zm00001d043131 | MYB转录因子 | Jiang et al., 2021 | | 10 | ZmMYB84 | Zm00001d025664 | MYB转录因子 | Jiang et al., 2021 | | 11 | ZmMs23 | Zm00001d008174 | bHLH转录因子 | Nan et al., 2017 | | 12 | ZmMs32 | Zm00001d006564 | bHLH转录因子 | Moon et al., 2013 | | 13 | ZmbHLH51 | Zm00001d053895 | bHLH转录因子 | Jiang et al., 2021 | | 14 | ZmbHLH122 | Zm00001d017724 | bHLH转录因子 | Jiang et al., 2021 | | 15 | ZmOCL4 | Zm00001d030069 | HD-ZIP转录因子 | Vernoud et al., 2009 | | 16 | ZmTGA9-1 | Zm00001d052543 | bZIP转录因子 | Jiang et al., 2021 | | 17 | ZmTGA9-2 | Zm00001d042777 | bZIP转录因子 | Jiang et al., 2021 | | 18 | ZmTGA9-3 | Zm00001d012294 | bZIP转录因子 | Jiang et al., 2021 | | 19 | ZmTGA10 | Zm00001d020938 | bZIP转录因子 | Jiang et al., 2021 | | 20 | ZmMs53 | Zm00001d053890 | SBP转录因子 | Liu et al., 2018 | | II 脂代谢类 | | 21 | ZmMs26 | Zm00001d027837 | 细胞色素P450单加氧酶, CYP704B1 | Djukanovic et al., 2013 | | 22 | ZmMs10/APV1 | Zm00001d024712 | 细胞色素P450单加氧酶, CYP703A2 | Somaratne et al., 2017 | | 23 | ZmMs30 | Zm00001d052403 | GDSL酯酶/脂肪酶 | An et al., 2019 | | 24 | ZmMs5/IPE2 | Zm00001d015960 | GDSL酯酶/脂肪酶 | Huo et al., 2020 | 25
| ZmMs33/GPAT6
| Zm00001d007714
| 甘油-3-磷酸酰基转移酶
| Xie et al., 2018; Zhang et al., 2018b; Zhu et al., 2020 | | 26 | ZmMs45 | Zm00001d047859 | 异胡豆苷合成酶 | Cigan et al., 2001 | | 27 | ZmMs20/IPE1* | Zm00001d029683 | GMC氧化还原酶 | Chen et al., 2017; Wang et al., 2019 | | 28 | ZmMs25/ZmMs6021 | Zm00001d048337 | 脂肪酰还原酶 | Tian et al., 2017; Zhang et al., 2021 | | 29 | ZmMs44 | Zm00001d052736 | 非特异性脂质转移蛋白 | Fox et al., 2017 | | 30 | ZmABCG26 | Zm00001d046537 | ABCG转运蛋白 | Jiang et al., 2021 | | 31 | ZmMs13 | Zm00001d013960 | ABCG转运蛋白 | Fang et al., 2023b | | 32 | ZmPKSB | Zm00001d019478 | 聚酮合酶B | Liu et al., 2022a | | 33 | ZmTKPR1-1 | Zm00001d031488 | 四肽α-吡咯酮还原酶 | An et al., 2024 | | 34 | ZmTKPR1-2 | Zm00001d020970 | 四肽α-吡咯酮还原酶 | An et al., 2024 | | III 糖代谢类 | | 35 | ZmMs8 | Zm00001d012234 | β-1,3-半乳糖基转移酶 | Wang et al., 2013 | | 36 | ZmMs39 | Zm00001d043909 | 胼胝质合成酶 | Zhu et al., 2018 | | 37 | ZmSTK1 | Zm00001d045056 | 丝氨酸苏氨酸激酶1 | Fan et al., 2018 | | 38 | ZmSTK2 | Zm00001d052067 | 丝氨酸苏氨酸激酶2 | Fan et al., 2018 | | 39 | ZmINVAN6/Mei025 | Zm00001d015094 | 胞质转化酶 | Huang et al., 2022 | | IV 其它途径类 | | 40 | ZmMs22/MSCA1 | Zm00001d018802 | 谷氧还蛋白 | Kelliher and Walbot, 2012 | | 41 | ZmMAC1 | Zm00001d023681 | 小分泌蛋白配体 | Wang et al., 2012 | | 42 | ZmMSP1 | Zm00001d042362 | 富亮氨酸重复受体激酶 | van der Linde et al., 2018 | | 43 | ZmDRP1 | Zm00001d035791 | 干燥相关蛋白 | Hu et al., 2022 | | 44 | ZmCOI2a | Zm00001d042833 | 茉莉酸受体 | Qi et al., 2022 | | 45 | ZmCOI2b | Zm00001d010082 | 茉莉酸受体 | Qi et al., 2022 | | 46 | ZmMs28/AGO5c | Zm00001d013063 | ARGONAUTE蛋白 | Li et al., 2021 | | 47 | ZmMs42/HSP101 | Zm00001d038806 | 热激蛋白 | Li et al., 2022 | | 48 | ZmTMS5 | Zm00001d053351 | RNA酶ZS1 | Li et al., 2017 | | 49 | ZmDCL5 | Zm00001d032655 | Dicer类似蛋白 | Teng et al., 2020 |
View table in article
表1
克隆鉴定的玉米核雄性不育(GMS)基因
正文中引用本图/表的段落
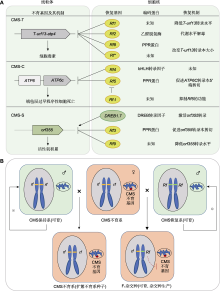
细胞质雄性不育(CMS)的不育性状受细胞质不育基因和细胞核育性恢复基因共同决定, 在玉米杂交种生产中应用最早(程计华等, 2006; 吴豪等, 2007; 郑名敏等, 2024)。根据育性恢复专效性, 玉米细胞质雄性不育材料可分为S型、C型和T型3种胞质类型(Beckett, 1971)。通过不育系与恢复系测交, 可以鉴定玉米细胞质雄性不育材料的胞质类型: 用待测不育材料作母本与恢复系恢313进行测交, 观察后代的育性表现, 育性恢复的材料为S型; 被保持的材料作为母本再与恢复系自凤1测交, 被恢复的材料为C型, 被保持的材料为T型(郑用琏, 1982)。T型和C型均属于孢子体不育, 育性受孢子体基因型决定, 败育发生在减数分裂期, 败育彻底, 育性较为稳定。S型属于配子体不育, 育性受花粉基因决定; 败育发生在减数分裂后的单核期与二核期, 多数不育材料出现育性不稳定现象(苏爱国等, 2018)。
玉米CMS-T型不育基因为线粒体基因T-urf13 (图1A), 该基因编码一个13 kDa的多肽(URF13)。URF13是一个线粒体内膜蛋白, 其通过影响线粒体内膜通透性导致电子传递链受损和能量代谢异常, 进而使花粉败育(Levings III, 1990)。CMS-T的育性恢复可由2个互补的显性核基因Rf1和Rf2控制。Rf1的恢复机制是促进T-urf13转录本加工, 产生2个大小分别为1.6 kb和0.6 kb的转录本, 进而抑制T-urf13的表达, 最终使URF13的蛋白丰度降低80%。Rf2基因编码一个玉米花药发育所必需的线粒体乙醛脱氢酶, 推测Rf2可能通过改变线粒体内的代谢水平来消除毒性蛋白URF13对线粒体的损害, 从而部分恢复T-urfl3导致的不育表型(Cui et al., 1996)。此外, Rf8和Rf*对影响不育基因T-urfl3的表达, 具有与Rf1相似但不完全相同的作用, 会产生更小的T-urf13转录本(Dill et al., 1997)。
玉米CMS-C型不育基因为线粒体基因atp6c (图1A)。与ATP6相比, ATP6C与ATP8和ATP9表现出更强的相互作用, 降低了F1F0-ATP合成酶的组装量和活性, 从而导致ATP产量降低、活性氧(ROS)爆发、绒毡层细胞过早程序性死亡和花粉败育(Yang et al., 2022)。CMS-C的育性恢复受Rf4和Rf5控制, 且具有重复效应。Rf4基因编码一个bHLH转录因子(Jaqueth et al., 2020), 目前其对CMS-C的育性恢复机制尚不清楚。Rf5基因编码一个P型PPR (pentatricopeptide repeat)蛋白, 通过招募一个剪接因子RS31A促进atp6c转录本的5'端部分剪切, 从而实现CMS-C的育性恢复(Lin et al., 2024)。此外, Rf5的育性恢复能力受Rf-I基因抑制, 具体作用机制还有待揭示(Hu et al., 2006)。
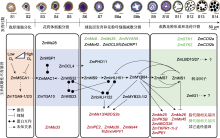
据不完全统计, 迄今玉米中采用正向(如图位克隆)和反向遗传学(如基因编辑)策略发掘的核不育(GMS)基因接近50个(表1)。这些基因按照其生物学功能或参与的代谢途径, 可分为转录因子、脂代谢、糖代谢和其它途径4种类型。其中转录因子20个, 包含LBD、PHD-finger、MYB、bHLH、HD-ZIP、bZIP和SBP共7类; 脂代谢基因14个, 包含脂合成和脂转运2类; 糖代谢基因5个; 其它类型基因10个。
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27。其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子。其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023)。ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007)。通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021)。
14个脂代谢类GMS基因中, 11个为脂合成相关基因, 即ZmMs5/IPE2、ZmMs6021/ZmMs25、ZmMs26、ZmMs30、ZmMs33、ZmMs20/IPE1、ZmMs10/APV1、ZmMs45、ZmTKPR1-1、ZmTKPR1-2和ZmPKSB; 另外3个为脂转运相关基因, 即ZmMs2/ZmABCG26、ZmMs13和ZmMs44 (表1)。
在玉米花药和花粉发育的脂质合成过程中, ZmMs6021/ZmMs25编码一个定位于质体的脂肪酰基还原酶, 催化脂肪酰基辅酶A (CoA)或酰基载体蛋白(ACP)底物还原为脂肪醇, 参与脂肪酸的从头合成, 这对于花药角质层的形成和花粉外壁发育至关重要(Tian et al., 2017; Zhang et al., 2021)。ZmMs26是拟南芥CYP704B1的同源基因, 编码一个细胞色素P450家族蛋白, 即长链脂肪酸ω-羟化酶, 参与孢粉素和某些生物大分子单体物质的合成(Djukanovic et al., 2013)。ZmMs30编码一个GDSL脂肪酶, 参与玉米花粉外壁足底层与花药角质层的发育(An et al., 2019)。ZmIPE2 (Irregular Pollen Exine2)也编码一个GDSL脂肪酶, ipe2突变体脂质代谢异常, 花药绒毡层细胞和中间层细胞不能正常降解, 最终导致花药角质层和花粉外壁发育异常(Huo et al., 2020)。ZmMs33编码甘油-3-磷酸酰基转移酶, 该基因影响花药内皮层叶绿体的功能, 最终影响育性(Xie et al., 2018; Zhu et al., 2020)。ZmMs45编码一个异胡豆苷合成酶类似蛋白, 其突变体的花粉外壁无法正常形成(Cigan et al., 2001)。ZmMs10/APV1 (Abnormal Pollen Vacuolation1)基因编码一个细胞色素P450脂肪酸羟化酶, 催化C12脂肪酸形成羟化的C12脂肪酸(Somaratne et al., 2017)。ZmMs20/ZmIPE1 (Irregular Pollen Exine1)编码一个葡萄糖甲醇胆碱(GMC)氧化还原酶, 影响C16/C18 ω-羟基脂肪酸的氧化过程, 与ZmMs26、ZmMs45和ZmAPV1共同调控花药角质层和花粉外壁的形成(Chen et al., 2017; Wang et al., 2019)。ZmPKSB编码一个III型聚酮合酶, 在体外能连续缩合脂酰辅酶A和丙二酰辅酶A产生四酮α-吡喃酮, pksb突变体表现为花粉外壁变薄, 但花药角质层增厚和变密。ZmTKPR1-1和ZmTKPR1-2编码四酮α-吡喃酮还原酶, 其对ZmPKSB产生的四酮a-吡咯酮具有还原活性, 同时ZmPKSB与ZmTKPR1- 1/-2互作, 形成控制花粉外壁形成的ZmPKSB-ZmTKPR1-1/-2孢粉素代谢区室(An et al., 2024)。
玉米花药和花粉发育过程中参与糖代谢的GMS基因有5个, 分别为ZmMs8、ZmMs39、ZmSTK1、ZmSTK2以及ZmINVAN6/Mei025 (表1)。ZmMs8编码一个β-1,3-半乳糖基转移酶, 其参与花药早期糖类的生物合成、绒毡层细胞程序性死亡和线粒体代谢等多个生物学过程; 该基因突变后, ms8突变体的花粉母细胞发育异常(Wang et al., 2013)。ZmMs39编码胼胝质合成酶ZmCalS12, 该基因在花药早期发育过程中参与胼胝质合成, ms39突变体的绒毡层细胞从二分体(S8a)时期开始出现异常, 最终表现为雄性不育及株高降低等性状(Zhu et al., 2018)。ZmSTK1和ZmSTK2编码受体样细胞质蛋白激酶, 调控玉米花粉发育并间接参与糖酵解途径; 它们突变均表现出严重的花粉传递缺陷, 从而影响花粉的育性(Fan et al., 2018)。玉米Mei025 (Meiotic025)是一个经典的显性核不育突变体, ZmMei025编码一个胞质转化酶INVAN6 (INVERTASE ALKALINE NEUTRAL 6), 主要在花粉母细胞中表达; INVAN6对于热胁迫下玉米花药的减数分裂必不可少, 该研究结果为通过操纵胞质糖代谢和信号转导来控制花粉发育的耐热性提供了新思路(Huang et al., 2022)。
已发现参与其它途径的玉米GMS基因有10个(表1)。其中, ZmMs28编码AGO (ARGONAUTE)家族蛋白ZmAGO5c, 该基因突变使绒毡层细胞在减数分裂早期提前出现空泡化, 其可能通过sRNA介导的表观遗传调控途径影响绒毡层细胞的发育(Li et al., 2021)。ZmMs22/MSCA1 (Male Sterile Converted Anther 1)编码一个CC型谷氧还蛋白, 参与调节孢原细胞的启动过程, ms22突变体中由于孢原细胞无法启动, 导致花药细胞分化成类似于叶片细胞的形态(Chaubal et al., 2003)。ZmMAC1 (Multiple Archesporial Cells 1)为拟南芥AtTPD1 (Tapetal Determinant 1)以及水稻OsTDL1A (TPD1-Like1A)的同源基因, 编码一个小分泌蛋白配体, 调控玉米花药发育早期孢原细胞的分裂增殖(Wang et al., 2012)。ZmMSP1 (Multiple Sporocyte 1)为拟南芥AtEMS1 (Excess Microsporocytes 1)和水稻OsMSP1的同源基因, 编码一个富亮氨酸重复受体样蛋白激酶(LRR-RLK)。与拟南芥和水稻的同源蛋白类似, ZmMSP1可与ZmMAC1直接互作, 形成一个功能保守的配体-受体调控模块(ZmMAC1-ZmMSP1), 促进花药发育早期孢原细胞的功能分化(van der Linde et al., 2018)。ZmCOI2a和ZmCOI2b编码F-box蛋白, 作为关键的茉莉酸(jasmonic acid, JA)受体, 冗余性地调控玉米花药开裂和配子体雄性育性; coi2a/coi2b双突变体的花粉在萌发和花粉管伸长方面均存在严重缺陷(Qi et al., 2022)。ZmDRP1 (Desiccation-Related Protein 1)基因在四分体到单核小孢子阶段的绒毡层和小孢子中特异性表达, ZmDRP1突变导致乌氏体异常、花粉外壁顶缺陷和完全雄性不育(Han et al., 2022; Hu et al., 2022)。
在孢原细胞特化后, 另一个重要的调节因子ZmMAC1被分泌出来, 阻止孢原细胞的进一步增殖, 并使邻近细胞成为支撑组织(Wang et al., 2012)。在孢原细胞外围的L2层细胞中, ZmMAC1被蛋白激酶ZmMSP1识别并与之相互作用, 促进L2层细胞分化为初生壁细胞(van der Linde et al., 2018)。当初生壁细胞分裂为内皮层细胞和次生壁细胞时, HD-ZIP IV转录因子ZmOCL4可抑制内皮层细胞的平周分裂。ocl4突变体花药内皮层和中间层之间多出一层具有内皮层特征的细胞层, 这导致部分雄性不育。ocl4/mac1双突变体表现出加性缺陷表型, 表明ZmOCL4和ZmMAC1通过不同途径独立作用, 但均调节表皮下的细胞增殖(Vernoud et al., 2009)。ZmMs23和ZmMs32作为bHLH转录因子能够互作, 共同调控绒毡层细胞的分裂和分化。ZmMs23转录本最初定位于次生壁前体细胞, 随后主要定位于其衍生的绒毡层细胞(Nan et al., 2017)。ZmMs32突变导致绒毡层前体细胞分化失败, 形成额外的绒毡层, 与野生型相比, ms32突变体中ZmMAC1的表达量降低, 这意味着ZmMAC1的表达需要ZmMS32 (Moon et al., 2013)。此外, 根据基因的表达时期, ZmTGA10、ZmMs33和ZmMs28可能也参与此生物学事件, 然而这些基因间的分子调控关系尚不清楚。
在玉米中已经鉴定了许多在花药发育后期(S9-S14)表达和发挥功能的GMS基因, 包括5个转录因子、8个脂质代谢相关基因和4个其它途径GMS基因(图2)。其中, PHD-finger类转录因子ZmMS7与玉米核因子(NF-Y)亚基相互作用形成ZmMS7-NF-YA6-YB2-YC9/ 12/15蛋白复合物, 通过直接结合下游靶基因启动子区域的CCAAT盒激活其下游靶基因的表达, 调控花粉外壁和花药角质层的形成(An et al., 2020)。最新研究表明, ZmMs1/ZmLBD30位于ZmbHLH51-ZmMYB84-ZmMS7的转录激活级联下游并被激活表达, 然后作为抑制因子通过反馈抑制关闭该级联激活通路, 通过直接抑制下游脂代谢GMS基因(如ZmMs26和ZmMs45), 或通过抑制ZmMYB84间接抑制下游脂代谢GMS基因(如ZmMs2/ABCG26、ZmTKPR1-1/2、ZmMs25、ZmPKSB、ZmIPE1、ZmMs26和ZmMs45), 以确保花粉发育过程中绒毡层及时退化、合适的孢粉素合成及花粉外壁厚度(Zhang et al., 2021; Liu et al., 2022a; Fang et al., 2023b; Hou et al., 2023)。基因表达和突变体表型分析表明, ZmIG1、ZmLBD10和ZmLBD27参与调控玉米花药发育后期进程, 但其分子机制尚不清楚(Jiang et al., 2021)。
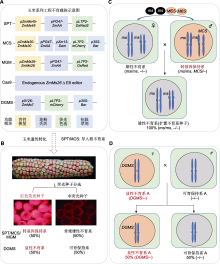
多数作物的雄配子对草甘膦非常敏感, 在合适的时期喷施草甘膦会导致植物雄性不育。据此, 研究人员创建了玉米草甘膦诱导型雄性不育技术。该技术通过组成型表达启动子35S驱动草甘膦不敏感基因CP4-EPSPS获得抗草甘膦玉米, 但在玉米花粉中CP4-EPSPS表达量很低, 同时优化草甘膦喷施参数, 即在V8及以后时期以单倍剂量(0.84 kg·hm-2)喷施玉米上部叶片, 使草甘膦高效积聚于雄穗中, 从而导致雄穗对草甘膦高度敏感并表现为草甘膦诱导型雄性不育。基于该技术, 美国孟山都公司首先创建了RHS1.0杂交制种系统(Feng et al., 2014)。2018年, 进一步建立了基于RNA干扰(RNAi)与CP4 EPSPS结合的第二代RHS (RHS2.0)技术。此技术利用玉米内源性雄性组织特异性小干扰RNA (mts-siRNA)触发雄穗中CP4 EPSPS mRNA的特异性裂解, 导致由于缺乏CP4 EPSPS蛋白而产生草甘膦敏感的雄性细胞; 然后在花粉发育的关键阶段使用草甘膦诱导雄性不育, 雄性不育的植株作为母本产生杂交种。该技术结合除草剂喷洒方法的相对简便以及靶向蛋白表达, 以不依赖于环境的方式创建了草甘膦诱导型雄性不育系统(Yang et al., 2018)。RHS系统的优势在于: (1) 一系两用, 不需要保持系; (2) 制种高效、种子纯度高且抗除草剂; (3) 不需要人工去雄, 降低了人工制种成本。RHS技术的不足在于喷施除草剂的窗口期较短, 具有潜在的母本不育性诱导不彻底风险。
目前, 玉米雄性不育相关研究取得了一些突破性进展, 在不育基因克隆与机制解析方面, 不断缩小与模式植物的差距, 已经完成约50个不育基因的克隆鉴定(表1), 同时利用现代前沿生物技术, 创建了多种玉米新型工程核不育技术体系, 如SPT、MCS、MGM和RHS技术, 且部分已实现产业化应用, 展示了工程核不育技术在玉米制种产业中的巨大价值。
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ... MALE STERILE6021 (MS6021) is required for the development of anther cuticle and pollen exine in maize 2 2017 ... The cloned genic male-sterility (GMS) genes in maize
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ... ZmMs25encoding a plastid-localized fatty acyl reductase is critical for anther and pollen development in maize 3 2021 ... The cloned genic male-sterility (GMS) genes in maize
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
20个转录因子类GMS基因中(表1), LBD转录因子有4个, 分别为ZmMs1、ZmIG1、ZmLBD10和ZmLBD27.其中ZmMs1编码绒毡层特异表达的LBD转录因子ZmLBD30, 为玉米花药和花粉发育中发现的第1个转录抑制因子.其可确保花粉发育过程中绒毡层的及时降解并精确调控花粉外壁形成(Hou et al., 2023).ZmIG1 (Indeterminate Gametophyte1)基因突变导致产生来自父系的单倍体胚囊和雄性不育(Evans, 2007).通过基因编辑产生的lbd10和lbd27单突变体育性正常, 而lbd10/27双突变体表现出部分雄性不育, 约32%的花粉粒呈现败育, 表明ZmLBD10和ZmLBD27影响育性并在花药发育中存在功能冗余(Jiang et al., 2021). ...
本文的其它图/表
|

 首页
首页