植物SPL转录因子的生物功能研究进展
曾鑫海, 陈锐, 师宇, 盖超越, 范凯, 李兆伟
植物学报
2023, 58 ( 6):
982-997.
DOI: 10.11983/CBB22216
SPL是植物特有的一类转录因子, 其蛋白结构中存在一段由2个锌指结构和核定位序列构成的高度保守的SBP结构域, 多数SPL基因的转录表达受microRNAs剪切调控。该文结合当前SPLs转录因子的研究进展, 对其在植物生长发育和环境适应等方面的生物学功能进行综述, 并对SPLs的研究前景进行展望。
| 物种 | 拉丁名 | SPL名称 | 功能 | 参考文献 | | 欧洲山杨 | Populus tremula | PtSPLs | MicroRNA156靶向调控SPL表达进而提高欧洲山杨花青素、黄酮和黄酮醇的生物合成 | Wang et al., 2020b | | 丹参 | Salvia miltiorrhiza | SmSPL6 | SmSPL6通过结合Sm4CL9和SmCYP98A14的启动子激活其表达, 促进SalB和RosA的生物合成 | Cao et al., 2021 | | SmSPL7 | SmSPL7通过直接与SmTAT1和Sm4CL9的启动子结合抑制其表达, 阻断SalB的生物合成 | Chen et al., 2021 | | 黄花蒿 | Artemisia annua | AaSPL2 | AaSPL2通过结合青蒿素合成关键基因DBR2的启动子促进其表达, 增强青蒿素的合成 | Lv et al., 2019 | | 广藿香 | Pogostemon cab-
lin | PaSPL9 | SPL9通过结合倍半萜合酶基因TPS21的启动子激活其表达, 促进老龄组织中倍半萜的生物合成 | Yu et al., 2015 | | 川桑 | Morus notabilis | MnSPL7 | MnSPL7通过激活儿茶素合成途径MnTT2L2基因的转录, 响应家蚕的取食行为 | Li et al., 2022a |
View table in article
表3
SPLs参与植物的次生代谢
正文中引用本图/表的段落
SPL转录因子能调控根系的分生区和根冠发育。Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性。Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控。此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控。
另外, SPL还参与拟南芥从营养生长向生殖生长的转换。Rahimi等(2022)研究发现, 拟南芥SPLs能通过互作阻遏AHL15的表达, 抑制营养生长, 促进植株从幼年营养生长向成年生殖生长转变(juvenile to adult vegetative phase change, VPC)以及开花。其中, 转录因子AtSPL3直接调控FUL (Fruitfull)以及AP1 (Ape-tala1)的转录, AtSPL4和AtSPL5能够促进FUL和SOC1 (Suppressor of overexpression of constans1)的转录, AtSPL9则正调控FUL、SOC1以及AGL42 (Agamous-like), 而FUL、SOC1、AGL42及AP1均促进植物开花(Birkenbihl et al., 2005; Wu and Poethig, 2006; Yamaguchi et al., 2009)。后续, Jung等(2012)揭示SOC1还能直接与AtSPL3、AtSPL4和AtSPL5 的启动子结合调控其表达。Yao等(2019b)的研究则发现, AtSPL10通过与MED25互作, 共同调控FUL和LFY (Leafy)等开花相关基因的表达, 从而促进过表达AtSPL10的拟南芥提前开花, 拟南芥atspl2/atspl10/atspl11三突变体则延迟开花。此外, Jiang等(2019)发现枇杷(Eriobotrya japonica)转录因子EjSPL4、EjSPL5和EjSPL9能够激活花发育关键基因EjSOC1-1、EjLFY-1和EjAP1-1的表达, 并且EjSPL3、EjSPL4、EjSPL5及EjSPL9在拟南芥中过表达均表现出花期提前现象, 表明EjSPL3、EjSPL4、EjSPL5和EjSPL9参与花期调控。柳枝稷中, 转录因子PvSPL7和PvSPL8通过调控SEP3 (Sepallata3)和MADS32的表达进而调节其开花时间(Gou et al., 2019)。过表达PvSPL6、PvSPL7和PvSPL8的转基因株系均较野生型更早开花, 而下调PvSPL6、PvSPL7和PvSPL8的表达, 可导致植株延迟开花, 甚至不开花(Gou et al., 2019; Cai et al., 2022)。Feng等(2023)通过对VcSPL序列进化关系、基因表达谱和启动子序列进行分析, 表明转录因子VcSPL35、VcSPL40、VcSPL45和VcSPL53在蓝莓(Vaccinium corymbosum)由营养生长向胚胎发生的过渡阶段发挥重要作用。Wang等(2020a)在牡丹(Paeonia × suffruticosa)研究中也发现, PsSPL2、PsSPL3、PsSPL9、PsSPL10、PsSPL13和PsSPL13A是调控牡丹成花转变的重要转录因子。
次生代谢是生物合成非必需物质并储存代谢产物的过程, 一些植物次生代谢产物具有一定的药用和工业价值。转录因子SPL也参与部分次生代谢过程, 其中以花青素的合成调控较多(表3)。花青素是一类重要的植物次生代谢物质, 经由黄酮类途径合成, 参与保护植物免受生物/非生物逆境胁迫。Wang等(2020b)的研究表明, microRNA156通过靶向调控SPL基因的mRNA表达水平, 促进欧洲山杨(Populus tremula)花青素、黄酮和黄酮醇的生物合成。在药用模式植物丹参(Salvia miltiorrhiza)中鉴定到15个SPL转录因子, 其中SmSPL6基因过表达可促进丹参中的酚酸积累, 抑制花青素合成, 且转基因丹参植株和根系中的主要水溶性有效成分丹酚酸(salvianolic acid B, SalB)和迷迭香酸(rosmarinic acid, RosA)含量均高于野生型, 通过酵母单杂交和双荧光素酶瞬时转录实验表明, 转录因子SmSPL6通过结合Sm4CL9和SmCYP98A14的启动子增强其表达, 进而促进SalB和RosA的生物合成(Cao et al., 2021)。Chen等(2021)研究表明, 丹参SmSPL7通过直接结合SmTAT1和Sm4CL9的启动子抑制其表达, 阻断SalB的生物合成, 并促进花青素积累。在青蒿素合成过程中, 转录因子AaSPL2通过结合青蒿素合成关键基因DBR2启动子的GTAC顺式作用元件增强其表达, 提高青蒿素的合成(Lv et al., 2019)。在广藿香(Pogostemon cablin)中, 转录因子SPL9通过直接结合倍半萜合酶基因TPS21的启动子激活其表达, 促进老龄组织中倍半萜的生物合成(Yu et al., 2015)。木本植物川桑(Morus notabilis)中, 转录因子MnSPL7通过激活儿茶素合成途径MnTT2L2基因的转录响应家蚕的取食行为(Li et al., 2022a)。
AtCBF2是拟南芥低温响应基因, 调控85个低温诱导基因和8个低温抑制基因的表达。当拟南芥遭受低温胁迫时, AtCBF2及下游低温响应基因被快速诱导表达(Vogel et al., 2005)。Zhao等(2022b)研究表明, 拟南芥转录因子AtSPL9通过直接与AtCBF2的启动子结合正调控其表达, 参与拟南芥对低温冻害的耐受性调节。高温胁迫下, 拟南芥AtSPL1和AtSPL12通过激活由ABA受体PYL (pyrabactin resistance 1-like)介导的ABA信号转导途径, 提高拟南芥花序的耐热性, 降低花器官对高温胁迫的敏感性(Chao et al., 2017)。当水稻遭受低温逆境时, OsmicroRNA156通过调控转录因子OsSPL3的mRNA水平, 降低OsSPL3的表达丰度, 减弱转录因子OsSPL3对OsWRKY71表达的正调控作用, 进而消除OsWRKY71对下游抗寒基因的负调控, 增强抗寒性(Zhou and Tang, 2019)。此外, Shan等(2021)在耐冷型甘蓝(Brassica oleracea var.capitata) CT-923中发现, 转录因子BoSPL9b和BoSPL16b在低温胁迫下的表达丰度显著上调; 在苜蓿中过表达microRNA156或者对MsSPL13进行靶向RNAi敲除突变, 能够使苜蓿对热胁迫的抗性增强, 表明了转录因子MsSPL13负调节苜蓿的耐热性(Matthews et al., 2019)。
转录因子SPL还参与植株对金属离子的吸收和利用。Garcia-Molina等(2014)研究发现, 定位于细胞内质网上的转录因子AtSPL7在胞内缺乏铜(Cu)时被激活, AtSPL7直接与Cu响应基因启动子上的响应元件(CuRE)结合, 促进铜还原酶基因FRO4/5和质膜转运蛋白COPT1/2/6表达, 进而增强对Cu的摄取(Yamasaki et al., 2009; Bernal et al., 2012)。此外, Perea- García等(2021)进一步揭示, AtSPL3过表达拟南芥抑制microRNA156对AtSPLs转录水平的调控, 减弱AtSPL7介导的Cu缺乏响应, AtSPL3可能参与对Cu响应基因启动子顺式作用元件的竞争性结合。Lu等(2011)通过系统发育分析表明, 毛果杨(Populus trichocarpa)转录因子PtSPL3和PtSPL4是CuRE结合蛋白, 调控Cu响应基因的表达, 但PtSPL3和PtSPL4的启动子序列中不存在CuRE, 其转录表达不受Cu操控。镉(Cd)胁迫下, 番茄LeSPL转录因子能直接结合SINR (Solanum lycopersicum nitrate reductase)的启动子抑制其表达, 从而负调控硝酸还原酶(nitrate reductase, NR)活性, 而NR能抑制Cd胁迫诱导的一氧化氮(nitric oxide, NO)生成(Chen et al., 2018)。
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
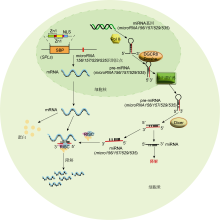
独脚金内酯(strigolactones, SLs)是高等植物中普遍存在的一种新型植物激素.近年来, 陆续有研究表明转录因子SPLs参与独脚金内酯信号转导与调控过程, 进而影响植物分蘖等表型构建.D53 (DWARF53)是SL信号通路的关键阻遏因子, Song等(2017)研究发现, 水稻D53能够与IPA1 (Ideal Plant Architecture1, 即转录因子OsSPL14)互作, 抑制IPA1的转录, 且IPA1还能直接与D53的启动子结合调控其表达.玉米中也发现有类似的调控途径.例如, Liu等(2021)指出, D53与UB3和TSH4存在互作, 并抑制它们对TB1的转录激活, 进而促进玉米分蘖, 且D53还可以结合UB3和TSH4的启动子抑制其转录.此外, Yang等(2022)研究发现, 柳枝稷PvSPL2可直接正调控SL生物合成重要基因PvLBO的表达. ... Transcriptome sequencing identifies SPL7-regulated copper acquisition genes FRO4/FRO5 and the copper dependence of iron homeostasis in Arabidopsis 1 2012 ... 转录因子SPL还参与植株对金属离子的吸收和利用.Garcia-Molina等(2014)研究发现, 定位于细胞内质网上的转录因子AtSPL7在胞内缺乏铜(Cu)时被激活, AtSPL7直接与Cu响应基因启动子上的响应元件(CuRE)结合, 促进铜还原酶基因FRO4/5和质膜转运蛋白COPT1/2/6表达, 进而增强对Cu的摄取(Yamasaki et al., 2009; Bernal et al., 2012).此外, Perea- García等(2021)进一步揭示, AtSPL3过表达拟南芥抑制microRNA156对AtSPLs转录水平的调控, 减弱AtSPL7介导的Cu缺乏响应, AtSPL3可能参与对Cu响应基因启动子顺式作用元件的竞争性结合.Lu等(2011)通过系统发育分析表明, 毛果杨(Populus trichocarpa)转录因子PtSPL3和PtSPL4是CuRE结合蛋白, 调控Cu响应基因的表达, 但PtSPL3和PtSPL4的启动子序列中不存在CuRE, 其转录表达不受Cu操控.镉(Cd)胁迫下, 番茄LeSPL转录因子能直接结合SINR (Solanum lycopersicum nitrate reductase)的启动子抑制其表达, 从而负调控硝酸还原酶(nitrate reductase, NR)活性, 而NR能抑制Cd胁迫诱导的一氧化氮(nitric oxide, NO)生成(Chen et al., 2018). ... Functional dissection of the plant-specific SBP-domain: overlap of the DNA-binding and nuclear localization domains 4 2005 ... SPL转录因子结构中包含一段约80个氨基酸残基的高度保守序列, 称为SBP结构域.核磁共振分析表明, SBP结构域含有2个典型的锌指结构, 包含8个半胱氨酸或组氨酸, 每4个氨基酸残基与1个锌离子结合, 分别形成Zn-1 (Cys-Cys-Cys-His或Cys-Cys- Cys-Cys)以及Zn-2 (Cys-Cys-His-Cys) (Yamasaki et al., 2004).SBP结构域的C端包含1个核定位信号 (nuclear localization signal, NLS)序列, 其与Zn-2结构序列存在部分重叠, 能引导SPL转录因子进入细胞核, 进而调控下游基因的表达.同时, SBP结构域能与含顺式作用元件TNCGTACAA的启动子结合, 其中GTAC为核心序列(Birkenbihl et al., 2005).此外, 部分SPL转录因子序列有1个高度保守的microRNA156识别位点, 少部分则含有microRNA529/535识别位点(Rhoades et al., 2002), 这是SPL基因转录后调控的主要方式. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ... Genomic organization, phylogenetic comparison and differential expression of the SBP-box family genes in grape 1 2013 ... 基因的表达调控可在多个水平上进行, 如转录、转录后、翻译及翻译后修饰.目前研究比较深入的是SPL转录因子在转录后水平的调控(图1).其中, 与microRNA156结合并降解SPL转录产生的mRNA为主要途径.microRNA156最初在拟南芥中发现, 是长度约为20个核苷酸的非编码单链RNA序列, 结构较为保守, 能特异性识别特定SPL转录产生的mRNA序列, 并通过剪切降解mRNA片段使其不能正常翻译(Jones- Rhoades et al., 2006).目前研究发现, 在拟南芥的17个SPL基因中, 有11个SPL序列含有microRNA-156识别位点, 在水稻(Oryza sativa)的19个SPL基因中, 有12个SPL序列具有microRNA156识别位点.此外, 高粱(Sorghum bicolor)、玉米(Zea mays)、黄瓜(Cucumis sativus)、柳枝稷(Panicum virgatum)和葡萄(Vitis vinifera)中分别有10、19、11、21和12个SPL基因包含microRNA156识别位点(Rhoades et al., 2002; Schwab et al., 2005; Bonnet et al., 2010; Dai and Zhao, 2011; Hou et al., 2013; Wu et al., 2016), 可见microRNA156在SPL基因表达中发挥重要调控作用.Gandikota等(2007)对拟南芥AtSPL3基因的microRNA156识别位点进行突变, 发现突变植株中AtSPL3的转录水平显著提高, 而microRNA156的含量与植物的生理年龄呈负相关, 降低microRNA156活性会导致植物出现早熟表型(Cheng et al., 2021).AHL蛋白(AT-hook motif nuclear localized)可以通过提高microRNA156/157的表达量抑制SPL的mRNA翻译(Rahimi et al., 2022).将番茄(Solanum lycopersicum)的SISPL13与35S-microRNA156a载体在瞬时表达系统中共表达时, SISPL13蛋白的含量下降至较低水平(Cui et al., 2020).水稻OsSPL14可调控水稻穗分枝, 当OsSPL14基因的microRNA156识别位点发生单碱基突变时, 水稻穗的分枝数显著增多(Miura et al., 2010; Jiao et al., 2010).在柑橘(Citrus reticulata)中, microRNA156/SPL模型调控柑橘愈伤组织的体细胞胚胎发生(Long et al., 2018).Yun等(2022)在大豆(Glycine max)中发现, microRNA156能直接结合并降解GmSPL9d的mRNA片段, 进而负调控大豆结瘤. ... The elite alleles of OsSPL4 regulate grain size and increase grain yield in rice 1 2021a ... 当前研究表明, SPL转录因子在禾谷类作物的产量形成及浆果类植物的果实发育过程中也发挥重要调控作用.以水稻产量性状为例, SPL转录因子主要调控水稻的穗分枝、穗粒数及粒型等.例如, 有研究表明转录因子OsSPL4、OsSPL13以及OsSPL16通过调控细胞分裂素水平来增加粒宽, 进而影响水稻的粒型 (Wang et al., 2012; Si et al., 2016; Hu et al., 2021a).在水稻分蘖、穗分枝及穗粒数等性状形成过程中, 转录因子OsSPL9可在幼穗分化早期直接激活花序分生组织特性基因RCN1 (rice TERMINAL FLOWER1/CENTRORADIALIS homolog)的表达, 进而调控水稻的穗分枝和穗粒数(Hu et al., 2021b); Yuan等(2019)研究表明, OsSPL18通过直接结合水稻直立密穗控制基因DEP1的启动子正调控其表达, 进而调节水稻穗型; 与野生型相比, osspl18突变体穗长、穗粒数以及粒宽等产量性状指标均减小, 但是分蘖数相对增多.Miura等(2010)研究发现, 转录因子OsSPL14通过促进水稻生殖生长期的穗分枝进而提高稻谷产量.此外, Zhang等(2021)研究表明, OsSPL12通过结合水稻粒宽关键基因GW5 (Grain width 5)的启动子负调控水稻粒宽; 粳稻中低活性OsSPL12可增加粒宽, 籼稻中高活性OsSPL12可降低粒宽.Gupta等(2023)利用CRISPR-Cas9技术编辑TaSPL13基因的microRNA156识别元件位点, 发现突变植株的籽粒大小和穗粒数均较野生型增加.Tripathi等(2020)发现二穗短柄草(Brachypodium distachyon) BdSBP9突变株的穗长显著变短. ... OsSPL9 regulates grain number and grain yield in rice 2 2021b ... The SPLs are involved in the regulation of plant growth and development
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ... miR156-targeted and nontargeted SBP-box transcription factors act in concert to secure male fertility in Arabidopsis 2 2010 ... The SPLs are involved in the regulation of plant growth and development
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ... Roles of transcription factor SQUAMOSA promoter binding protein-like gene family in papaya (Carica papaya) development and ripening 2 2020 ... 激素在植物生长发育过程中发挥重要调节作用.SPL参与脱落酸(abscisic acid, ABA)、生长素(IAA)、细胞分裂素、赤霉素(gibberellin, GA)以及乙烯(ethylene)等激素的信号转导途径(表2).Dong等(2021)研究发现, 拟南芥AtSPL9通过直接结合ABA信号途径关键基因ABI5 (Abscisic Acid Insensitive 5)的启动子激活其表达, 促进ABA信号转导和种子中的ABA积累, 抑制拟南芥种子成熟后荚内发芽, 且AtSPL9受到蛋白激酶SnRK2的磷酸化调控.Bencivenga等(2012)研究拟南芥胚珠发育时, 发现转录因子AtSPL10和BEL1通过与生长素转运基因PIN1相互作用, 调节胚珠内的生长素与细胞分裂素分布, 从而调控胚珠发育.水稻转录因子OsSPL14通过激活生长素运输基因OsPIN1b以及PILS6b的表达参与生长素的极性运输(Li et al., 2022c).Qin等(2020)研究表明, 水稻OsSPL12转录因子通过与9个GA信号途径相关基因直接互作, 参与调控籽粒内源GA水平, 促进成熟籽粒休眠, 显著抑制稻谷收获前的穗发芽.Zhao等(2022a)以玉米株高调控基因D1 (dwarfplant1)为切入点, 研究发现转录因子ZmSPL12能直接与D1的启动子特异性结合抑制其转录, 降低玉米节间活性赤霉素含量, 从而抑制细胞伸长, 使玉米节间缩短, 株高降低, 有效提高玉米的抗倒伏能力.此外, Xu等(2020)研究表明, 在1-甲基环丙烯(1-methylcyclopropene, 1-MCP)处理下, 番木瓜(Carica papaya)中CpSPL3和CpSPL6随着果实不断成熟而上调表达, CpSPL11则在乙烯利(ethe-phon, ETH)处理的第8天表达水平明显高于未处理和1-MCP处理组, 并且CpSPL3、CpSPL6和CpSPL11在存储期具有与CpmicroRNA156相反的ETH/I-MCP响应模式, 表明CpmicroRNA156/CpSPL3/CpSPL6/ CpSPL11可能通过响应ETH/I-MCP信号参与番木瓜着色和成熟调节. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等(2020)研究发现, 当拟南芥AtSPL10表达受到抑制时, 可以通过调节细胞分裂素(cytokinin, CTK)途径来调控根系分生组织的活性.Shao等(2019)研究表明, 水稻OsSPL3和OsSPL12参与植株根冠生长的调节过程, 其中, OsSPL3与靶基因OsMADS50相互作用, 调节生长素的运输和信号转导, 进而影响根冠发育, 并且OsSPL3的表达受microRNA156调控.此外, microRNA156-OsSPL3-OsMADS50互作途径也参与水稻的不定根发育调控. ...
本文的其它图/表
|

 首页
首页