|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
植物SPL转录因子的生物功能研究进展
植物学报
2023, 58 (6):
982-997.
DOI: 10.11983/CBB22216
SPL是植物特有的一类转录因子, 其蛋白结构中存在一段由2个锌指结构和核定位序列构成的高度保守的SBP结构域, 多数SPL基因的转录表达受microRNAs剪切调控。该文结合当前SPLs转录因子的研究进展, 对其在植物生长发育和环境适应等方面的生物学功能进行综述, 并对SPLs的研究前景进行展望。
表4
SPLs参与植物的逆境胁迫响应
正文中引用本图/表的段落
当前研究表明, SPL转录因子在禾谷类作物的产量形成及浆果类植物的果实发育过程中也发挥重要调控作用。以水稻产量性状为例, SPL转录因子主要调控水稻的穗分枝、穗粒数及粒型等。例如, 有研究表明转录因子OsSPL4、OsSPL13以及OsSPL16通过调控细胞分裂素水平来增加粒宽, 进而影响水稻的粒型 (Wang et al., 2012; Si et al., 2016; Hu et al., 2021a)。在水稻分蘖、穗分枝及穗粒数等性状形成过程中, 转录因子OsSPL9可在幼穗分化早期直接激活花序分生组织特性基因RCN1 (rice TERMINAL FLOWER1/CENTRORADIALIS homolog)的表达, 进而调控水稻的穗分枝和穗粒数(Hu et al., 2021b); Yuan等(2019)研究表明, OsSPL18通过直接结合水稻直立密穗控制基因DEP1的启动子正调控其表达, 进而调节水稻穗型; 与野生型相比, osspl18突变体穗长、穗粒数以及粒宽等产量性状指标均减小, 但是分蘖数相对增多。Miura等(2010)研究发现, 转录因子OsSPL14通过促进水稻生殖生长期的穗分枝进而提高稻谷产量。此外, Zhang等(2021)研究表明, OsSPL12通过结合水稻粒宽关键基因GW5 (Grain width 5)的启动子负调控水稻粒宽; 粳稻中低活性OsSPL12可增加粒宽, 籼稻中高活性OsSPL12可降低粒宽。Gupta等(2023)利用CRISPR-Cas9技术编辑TaSPL13基因的microRNA156识别元件位点, 发现突变植株的籽粒大小和穗粒数均较野生型增加。Tripathi等(2020)发现二穗短柄草(Brachypodium distachyon) BdSBP9突变株的穗长显著变短。
利用基因芯片和miRNA测序技术, 研究人员发现植物中许多microRNAs响应生物/非生物逆境胁迫。转录因子SPL的表达受microRNAs的转录水平调控, 在植物响应生物和非生物胁迫中发挥重要调节作用(表4) (Sunkar and Zhu, 2004)。
在干旱或盐胁迫下, 植株通常呈现出生理性缺水现象, 组织细胞通过调节脯氨酸和可溶性糖等渗透物质含量, 并合成次生代谢物, 提高植株的保水能力和抗逆性。IPA1与SNAC1的启动子结合, 直接激活其表达, 从而调节ROS (reactive oxygen species)稳态, 赋予水稻植株耐旱性(Chen et al., 2023)。苜蓿抗旱研究表明, 转录因子MsSPL13参与苜蓿的耐旱性调节, 适度的microRNA156转录丰度可有效抑制MsSPL13的表达, 促进WD40-1的表达, 进而微调控DFR (Dihydroflavonol-4-reductase)的表达, 增强花青素的生物合成, 促进脯氨酸和可溶性糖积累, 从而提高苜蓿的抗旱性(Feyissa et al., 2019)。Hanly等(2020)研究发现, 转录因子MsSPL9的RNAi沉默转基因苜蓿比野生型苜蓿更能够适应干旱环境。对木薯(Manihot esculenta)的研究表明, 转录因子MeSPL9通过调控JA信号和抗氧化物质水平来调节木薯的抗旱能力(Li et al., 2022b)。此外, Wang等(2021b)研究发现, 美洲山核桃(Carya illinoinensis) CiSPL基因家族在干旱和盐胁迫下呈现出不同的时空表达特性, 以应对逆境胁迫。
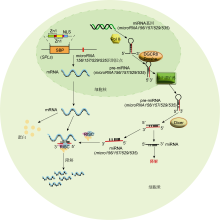
独脚金内酯(strigolactones, SLs)是高等植物中普遍存在的一种新型植物激素.近年来, 陆续有研究表明转录因子SPLs参与独脚金内酯信号转导与调控过程, 进而影响植物分蘖等表型构建.D53 (DWARF53)是SL信号通路的关键阻遏因子, Song等( Transcriptome sequencing identifies SPL7-regulated copper acquisition genes FRO4/FRO5 and the copper dependence of iron homeostasis in Arabidopsis 1 2012 ... 转录因子SPL还参与植株对金属离子的吸收和利用.Garcia-Molina等( Functional dissection of the plant-specific SBP-domain: overlap of the DNA-binding and nuclear localization domains 4 2005 ... SPL转录因子结构中包含一段约80个氨基酸残基的高度保守序列, 称为SBP结构域.核磁共振分析表明, SBP结构域含有2个典型的锌指结构, 包含8个半胱氨酸或组氨酸, 每4个氨基酸残基与1个锌离子结合, 分别形成Zn-1 (Cys-Cys-Cys-His或Cys-Cys- Cys-Cys)以及Zn-2 (Cys-Cys-His-Cys) (Yamasaki et al.,
利用基因芯片和miRNA测序技术, 研究人员发现植物中许多microRNAs响应生物/非生物逆境胁迫.转录因子SPL的表达受microRNAs的转录水平调控, 在植物响应生物和非生物胁迫中发挥重要调节作用( Functional analysis of the Arabidopsis thaliana SBP-box gene SPL3: a novel gene involved in the floral transition 1 1997 ... SPL (SQUAMOSA promoter binding protein- like)是一类广泛存在于绿色植物中的转录因子(transcription factor).1992年, Huijser等( Arabidopsis transcription factors SPL1 and SPL12 confer plant thermotolerance at reproductive stage 2 2017 ... SPLs participate in the response to various stresses
利用基因芯片和miRNA测序技术, 研究人员发现植物中许多microRNAs响应生物/非生物逆境胁迫.转录因子SPL的表达受microRNAs的转录水平调控, 在植物响应生物和非生物胁迫中发挥重要调节作用( LeSPL-CNR negatively regulates Cd acquisition through repressing nitrate reductase-mediated nitric oxide production in tomato 2 2018 ... SPLs participate in the response to various stresses
SPL转录因子能调控根系的分生区和根冠发育.Barrera-Rojas等( Genomic organization, phylogenetic comparison and differential expression of the SBP-box family genes in grape 1 2013 ... 基因的表达调控可在多个水平上进行, 如转录、转录后、翻译及翻译后修饰.目前研究比较深入的是SPL转录因子在转录后水平的调控( The elite alleles of OsSPL4 regulate grain size and increase grain yield in rice 1 2021a ... 当前研究表明, SPL转录因子在禾谷类作物的产量形成及浆果类植物的果实发育过程中也发挥重要调控作用.以水稻产量性状为例, SPL转录因子主要调控水稻的穗分枝、穗粒数及粒型等.例如, 有研究表明转录因子OsSPL4、OsSPL13以及OsSPL16通过调控细胞分裂素水平来增加粒宽, 进而影响水稻的粒型 (Wang et al., OsSPL9 regulates grain number and grain yield in rice 2 2021b ... The SPLs are involved in the regulation of plant growth and development
利用基因芯片和miRNA测序技术, 研究人员发现植物中许多microRNAs响应生物/非生物逆境胁迫.转录因子SPL的表达受microRNAs的转录水平调控, 在植物响应生物和非生物胁迫中发挥重要调节作用( TaSPL13 regulates inflorescence architecture and development in transgenic wheat (Triticum aestivum L.) 2 2020 ... The SPLs are involved in the regulation of plant growth and development
利用基因芯片和miRNA测序技术, 研究人员发现植物中许多microRNAs响应生物/非生物逆境胁迫.转录因子SPL的表达受microRNAs的转录水平调控, 在植物响应生物和非生物胁迫中发挥重要调节作用( Characterization of Squamosa-promoter binding protein-box family genes reveals the critical role of MsSPL20 in alfalfa flowering time regulation 2 2022 ... The SPLs are involved in the regulation of plant growth and development
利用基因芯片和miRNA测序技术, 研究人员发现植物中许多microRNAs响应生物/非生物逆境胁迫.转录因子SPL的表达受microRNAs的转录水平调控, 在植物响应生物和非生物胁迫中发挥重要调节作用( The molecular mechanism of SPOROCYTELESS/NOZZLE in controlling Arabidopsis ovule development 2 2015 ... The SPLs are involved in the regulation of plant growth and development
利用基因芯片和miRNA测序技术, 研究人员发现植物中许多microRNAs响应生物/非生物逆境胁迫.转录因子SPL的表达受microRNAs的转录水平调控, 在植物响应生物和非生物胁迫中发挥重要调节作用( OsSPL18 controls grain weight and grain number in rice 2 2019 ... The SPLs are involved in the regulation of plant growth and development
本文的其它图/表
|
 首页
首页