
|
|
||
|
植物抗冻蛋白研究进展
植物学报
2023, 58 (6):
966-981.
DOI: 10.11983/CBB22248
温度是决定植物地理分布的主要环境因子之一, 分布于高纬度、高海拔地区的寒地植物, 在其生活史周期内通常会遭遇一段时期的零度以下低温。当环境温度降至冰点以下, 植物体内水分子趋向于形成冰晶体, 不断增大的冰晶对植物组织结构造成致命损伤。为适应冷冻环境, 寒地植物病程相关蛋白(PR)及相关的WRKY转录因子进化成为能与冰面特异吸附结合、高效抑制冰晶形成和生长的抗冻蛋白(AFPs)。目前, 已从冬黑麦(Secale cereale)等近百种植物中分离鉴定了AFPs。与昆虫AFPs相比, 植物AFPs具有极高的重结晶抑制活性, 可有效防止形成体内大冰晶。低温和病原菌均可诱导寒地植物合成AFP。有趣的是, 仅冷诱导合成的AFPs有水解酶/抗冻活性双重分子功能。然而, 越来越多的证据表明, PR-AFP仅具有水解酶/抗冻活性的其中一种, 其转化由翻译后肽差异折叠控制/调节。AFP因具有独特的分子功能与广阔的应用前景而逐渐成为植物学领域的研究热点。该文对近年来相关领域取得的研究进展进行系统综述。 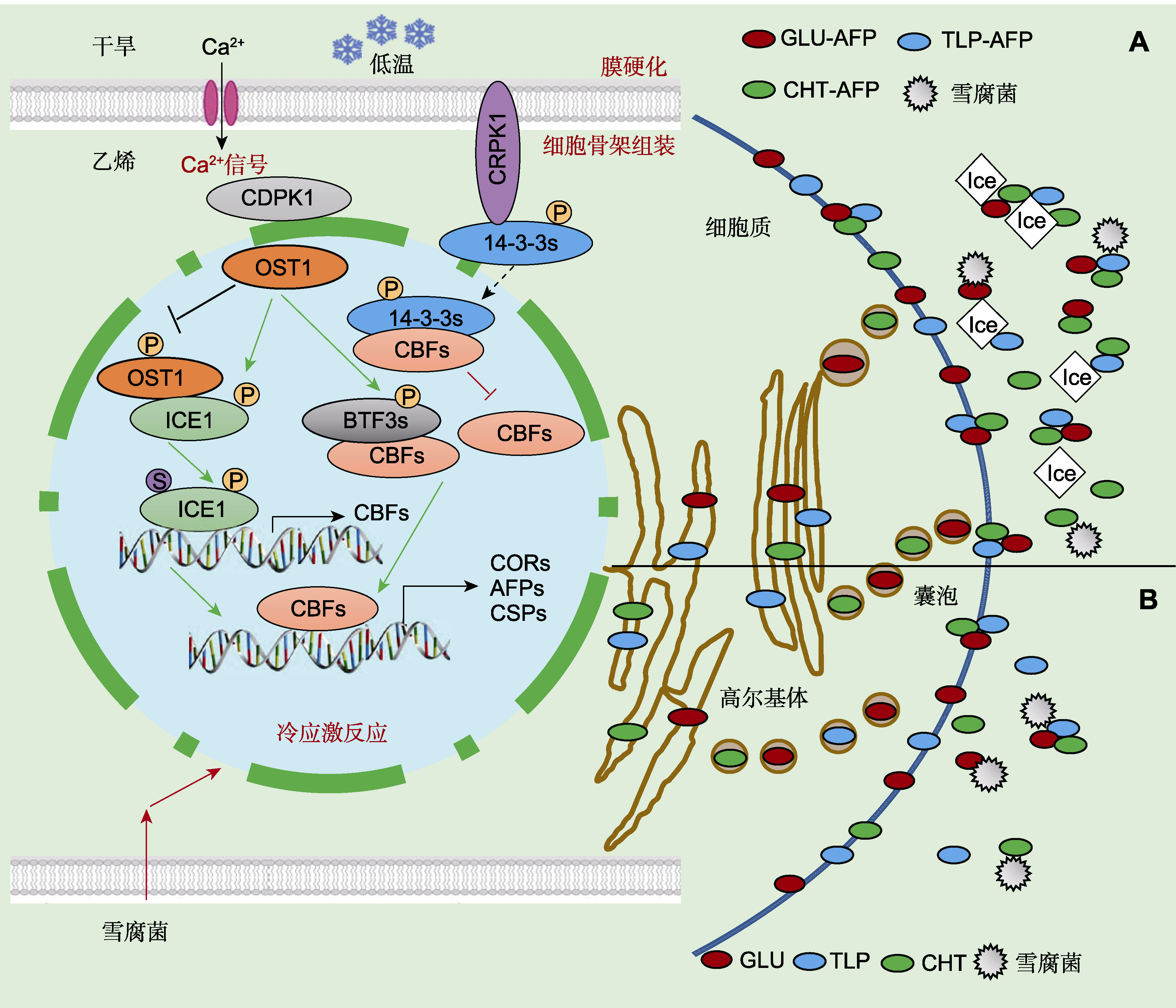 View image in article
图3
低温下植物抗冻蛋白(AFPs)和致病相关蛋白(PRs)的分子调控模型(Griffith and Yaish,
(A) 低温胁迫诱导COR、AFP和CSP等基因表达, 新合成的PR-AFP蛋白和PR蛋白通过内质网、高尔基体和囊泡分泌运输, 穿过质膜并聚集在细胞壁外空间, 形成寡聚体复合物, 抑制低温下病原菌生长; 这些蛋白质包括几丁质酶-AFPs (CHT-AFP)、葡聚糖酶-AFPs (GLU-AFP)和类甜蛋白-AFPs (TLP-AFP); 当环境温度降至冰点以下, PR-AFPs蛋白可特异吸附到冰晶表面, 抑制冰晶生长和重结晶; (B) PR蛋白(包括几丁质酶(CHT)、葡聚糖酶(GLU)和类甜蛋白(TLP))被合成并分泌到质外体中, 常温下用水杨酸(SA)、脱落酸(ABA)或雪腐病原菌处理植物, 仅诱导合成PR蛋白, 可抑制真菌病原体生长。
正文中引用本图/表的段落
植物细胞通过冷信号通路感知并应答环境温度, 环境冷信号被细胞质膜受体感知, 并转化为生物化学信号进入细胞内。不同植物感受冷信号的受体不同, 在单子叶植物水稻中COLD1可能是冷信号受体, COLD1编码G蛋白信号调控因子, 该因子定位于质膜和内质网, 可与G蛋白α亚基互作并激活质膜Ca2+通道。在双子叶植物拟南芥(Arabidopsis thaliana)中, 低温导致质膜状态发生变化, 引起G蛋白活化或直接活化质膜Ca2+通道蛋白CNGC; 质膜通道蛋白活化后Ca2+从细胞壁内流进入细胞, 作为第二信使通过CRPK1激酶和MAP3K-MKK4/5级联途径, 分别磷酸化激活14-3- 3s和MPK3/6, 活化的14-3-3s和MPK3/6穿过核膜从胞质进入核内, 分别参与CBF (C-repeat binding factors)和ICE1的泛素化降解(Yu et al., 2001)。ICE1- CBFs-COR通路处于冷信号转导网络的中心位置。CBF是一个转录因子, 可结合低温胁迫响应基因COR (cold-regulated)启动子顺式作用元件, 激活COR基因表达(Shi et al., 2015; 刘静妍等, 2017; Gao et al., 2018; Guo et al., 2018)。合成的PR-AFPs和PR通过内质网运输到高尔基体形成囊泡, 包裹蛋白的囊泡与质膜融合, 并将PR-AFPs或PR蛋白分泌释放到胞外壁表面(Pihakaski-Maunsbach et al., 1996)。当环境降温和/或受到病原菌侵染(Yu and Griffith, 1999; Hiilovaara-Teijo et al., 1999), PR-AFPs或PR蛋白可特异吸附到冰晶和病原菌上, 防止冰晶生长和病原菌侵染(图3A)。除环境冷信号诱导植物AFPs表达外, 常温下乙烯、茉莉酸和干旱可诱导合成PR-AFPs (Dave and Mitra, 1998), 而水杨酸、脱落酸和雪腐病原菌仅诱导合成PR (Hiilovaara-Teijo et al., 1999; 卢存福等, 2000; Yu and Griffith, 2001; Yu et al., 2001) (图3B)。
TH: 热滞活性; RI: 重结晶抑制活性 ... Psychrophilic lifestyles: mechanisms of adaptation and biotechnological tools 1 2019 ... AFPs-冰晶结合力的调节是AFPs抗冻机制的关键, 也是理解AFPs结构与功能的重点.早期研究表明, 氢键是AFPs与冰晶结合的作用力, AFPI亲水性残基Thr和Asp在二级螺旋上呈规律性间隔, Thr羟基与Asp羧基间的距离为4.5?, 这与冰晶平面上氧原子的间距正好匹配, 两者之间形成氢键, 从而使AFPs稳定结合冰面(DeVries, A low temperature induced apoplastic protein isolated from Arachis hypogaea 1 1998 ... 植物细胞通过冷信号通路感知并应答环境温度, 环境冷信号被细胞质膜受体感知, 并转化为生物化学信号进入细胞内.不同植物感受冷信号的受体不同, 在单子叶植物水稻中COLD1可能是冷信号受体, COLD1编码G蛋白信号调控因子, 该因子定位于质膜和内质网, 可与G蛋白α亚基互作并激活质膜Ca2+通道.在双子叶植物拟南芥(Arabidopsis thaliana)中, 低温导致质膜状态发生变化, 引起G蛋白活化或直接活化质膜Ca2+通道蛋白CNGC; 质膜通道蛋白活化后Ca2+从细胞壁内流进入细胞, 作为第二信使通过CRPK1激酶和MAP3K-MKK4/5级联途径, 分别磷酸化激活14-3- 3s和MPK3/6, 活化的14-3-3s和MPK3/6穿过核膜从胞质进入核内, 分别参与CBF (C-repeat binding factors)和ICE1的泛素化降解(Yu et al., Ice-binding proteins: a remarkable diversity of structures for stopping and starting ice growth 1 2014 ... 生长的冰晶将接近的溶质从冰面附近推开, 使水分子不断进入到晶格中, 导致冰晶体增大.而AFPs与冰晶具有特殊的亲和力, 能非常稳定地结合在冰晶表面.冰晶表面以直平面的形式向前伸展生长, AFPs结合冰面后产生一个小于正常冰晶半径的曲率半径, 这种从直平面到曲面的改变导致与水接触的冰表面积和表面张力增大, 从而使更多的能量被释放; 冰晶表面积的增大使体系的平衡状态发生改变, 从而使整个体系的冰点降低, 产生热滞效应(Davies, Antifreeze peptides and glycopeptides in cold-water fishes 1 1983 ... AFPs-冰晶结合力的调节是AFPs抗冻机制的关键, 也是理解AFPs结构与功能的重点.早期研究表明, 氢键是AFPs与冰晶结合的作用力, AFPI亲水性残基Thr和Asp在二级螺旋上呈规律性间隔, Thr羟基与Asp羧基间的距离为4.5?, 这与冰晶平面上氧原子的间距正好匹配, 两者之间形成氢键, 从而使AFPs稳定结合冰面(DeVries, Antifreeze glycopeptides and peptides: interactions with ice and water 2 1986 ... 植物AFPs亲和吸附在冰晶表面, 可降低细胞溶液的冰点, 而其熔点几乎不变, 这种性质称为热滞活性(TH) (DeVries,
(A) Cold stress induces COR, AFP and CSP gene expression; newly synthesized, dual-function PR-AFP proteins and PRs are secreted via the endoplasmic reticulum, Golgi bodies and vesicles that merge with the plasmalemma and accumulate on the outer cell wall surface; the AFPs, including chitinase-AFPs (CHT-AFP), glucanase-AFPs (GLU-AFP) and thaumatin-like AFPs (TLP-AFP), form oligomeric complexes that inhibit the growth of pathogens under low temperature; Below the freezing degree, the proteins are adsorbed onto the surfaces of ice and inhibit its growth and recrystallization; (B) PR proteins, including chitinases (CHT), glucanases (GLU), and thaumatin-like proteins (TLP), were synthesized and secreted into the apoplast, when plants are treated with salicylic acid (SA), abscisic acid (ABA) or snow mold at normal temperatures to inhibit the growth of fungal pathogens. ... Ice recrystallization inhibition activity varies with ice-binding protein type and does not correlate with thermal hysteresis 1 2021 ... 当环境温度降到冰点以下时, 植物细胞溶液中先形成细小晶核, 晶核相互融合长大, 冰晶体增大会对植物组织和细胞产生严重的物理损伤(Griffith et al., Cold signaling in plants: insights into mechanisms and regulation 3 2018 ... 植物细胞通过冷信号通路感知并应答环境温度, 环境冷信号被细胞质膜受体感知, 并转化为生物化学信号进入细胞内.不同植物感受冷信号的受体不同, 在单子叶植物水稻中COLD1可能是冷信号受体, COLD1编码G蛋白信号调控因子, 该因子定位于质膜和内质网, 可与G蛋白α亚基互作并激活质膜Ca2+通道.在双子叶植物拟南芥(Arabidopsis thaliana)中, 低温导致质膜状态发生变化, 引起G蛋白活化或直接活化质膜Ca2+通道蛋白CNGC; 质膜通道蛋白活化后Ca2+从细胞壁内流进入细胞, 作为第二信使通过CRPK1激酶和MAP3K-MKK4/5级联途径, 分别磷酸化激活14-3- 3s和MPK3/6, 活化的14-3-3s和MPK3/6穿过核膜从胞质进入核内, 分别参与CBF (C-repeat binding factors)和ICE1的泛素化降解(Yu et al.,
TH: 热滞活性; RI: 重结晶抑制活性 ... Computationally efficient approach for the identification of ice-binding surfaces and how they bind ice 1 2020 ... AFPs的冰晶结合位点所形成的表面与冰晶表面形成互补, 相互作用力主要是疏水作用和范德华力, 氢键起次要作用(Naullage et al., The effects of antifreeze proteins on chilled and frozen meat 1 1994 ... AFPs可与冰晶表面特异亲和吸附, 降低细胞溶液的冰点, 在冷冻和解冻过程中抑制冰晶重结晶(Yeh and Feeney, Immunogold localization of glucanase-like antifreeze protein in cold acclimated winter rye 1 1996 ... 植物细胞通过冷信号通路感知并应答环境温度, 环境冷信号被细胞质膜受体感知, 并转化为生物化学信号进入细胞内.不同植物感受冷信号的受体不同, 在单子叶植物水稻中COLD1可能是冷信号受体, COLD1编码G蛋白信号调控因子, 该因子定位于质膜和内质网, 可与G蛋白α亚基互作并激活质膜Ca2+通道.在双子叶植物拟南芥(Arabidopsis thaliana)中, 低温导致质膜状态发生变化, 引起G蛋白活化或直接活化质膜Ca2+通道蛋白CNGC; 质膜通道蛋白活化后Ca2+从细胞壁内流进入细胞, 作为第二信使通过CRPK1激酶和MAP3K-MKK4/5级联途径, 分别磷酸化激活14-3- 3s和MPK3/6, 活化的14-3-3s和MPK3/6穿过核膜从胞质进入核内, 分别参与CBF (C-repeat binding factors)和ICE1的泛素化降解(Yu et al., Changes in activities of antioxidant enzymes and their relationship to genetic and paclobutrazol-induced chilling tolerance of maize seedlings 2 1997 ... 温度是决定植物地理分布的主要环境因子之一(Yan et al.,
TH: 热滞活性; RI: 重结晶抑制活性 ... Antifreeze proteins in winter rye leaves form oligomeric complexes 1 1999 ... 植物细胞通过冷信号通路感知并应答环境温度, 环境冷信号被细胞质膜受体感知, 并转化为生物化学信号进入细胞内.不同植物感受冷信号的受体不同, 在单子叶植物水稻中COLD1可能是冷信号受体, COLD1编码G蛋白信号调控因子, 该因子定位于质膜和内质网, 可与G蛋白α亚基互作并激活质膜Ca2+通道.在双子叶植物拟南芥(Arabidopsis thaliana)中, 低温导致质膜状态发生变化, 引起G蛋白活化或直接活化质膜Ca2+通道蛋白CNGC; 质膜通道蛋白活化后Ca2+从细胞壁内流进入细胞, 作为第二信使通过CRPK1激酶和MAP3K-MKK4/5级联途径, 分别磷酸化激活14-3- 3s和MPK3/6, 活化的14-3-3s和MPK3/6穿过核膜从胞质进入核内, 分别参与CBF (C-repeat binding factors)和ICE1的泛素化降解(Yu et al., Winter rye antifreeze activity increases in response to cold and drought, but not abscisic acid 2 2001 ... 在严寒环境中, 植物合成多种类型AFPs应对冷胁迫.例如, 低温环境下冬黑麦叶片质外体提取液具有抗冻活性, 已鉴定出6种抗冻蛋白, 包括2种葡聚糖酶(β-1,3-或β-1,4-葡聚糖酶)、2种几丁质酶和2种类甜蛋白(Hon et al.,
本文的其它图/表
|
 首页
首页

{kind=link}