|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
茶树茎段不定芽高效发生体系的建立
植物学报
2023, 58 (2):
308-315.
DOI: 10.11983/CBB22184
茶树(Camellia sinensis)是重要的经济作物, 杂合度高且变异度大, 其高效离体再生体系鲜见报道。以舒茶早茎段为起始外植体, 进行不定芽高效发生影响因子研究。结果表明, MS+2 mg∙L-1 6-BA为定芽诱导的最适配方, 定芽诱导率为84.44%, 吸收底盘膨大率为80%, 利于后续不定芽诱导; MS+2 mg∙L-1 6-BA+0.2 mg∙L-1 NAA+0.1 mg∙L-1 KT+1 mg∙L-1脯氨酸为不定芽增殖诱导的适宜配方, 不定芽诱导率为88.89%, 平均芽数为7.8。不定根诱导的适宜配方为1/2MS+3 mg∙L-1 IBA, 生根率为85.56%。采用RAPD和ISSR技术对再生植株进行分子检测, 在连续2代离体再生植株中未发现明显变异。
表6
随机扩增多态性DNA (RAPD)引物及其扩增谱
正文中引用本图/表的段落
为了筛选出茶树离体再生的最适基本培养基, 将舒茶早带叶柄和部分叶片的茎段(图1A)分别接种到添加1 mg?L-1 6-BA和0.1 mg?L-1 NAA的MS、WPM、DCR和N6四种基本培养基上, 进行茶树定芽诱导, 观察培养物的生长状况。结果表明, MS培养基中茎段的长势较好, 芽苗幼嫩, 叶片富有光泽, 整个培养物生长活力强; 而WPM培养基中的茎段长势较弱, 长出的芽茎较细, 叶片偏红; 在DCR培养基中长出的芽细瘦, 叶片偏黄; N6培养基中的叶柄稍微带红, 新芽瘦弱(表1)。因此, 确定MS培养基为茶树茎段定芽诱导的适宜基本培养基。不同基本培养基诱导茶树茎段定芽存在差异, 这可能与不同基本培养基中所含元素浓度不同有关。而MS培养基含有较高浓度的无机盐, N和K等元素含量也较高, 有利于定芽诱导。
为探明6-BA对定芽诱导的影响, 将带有叶柄和部分叶片的茎段接种到含不同浓度6-BA的MS培养基上进行定芽诱导, 4周后统计定芽诱导率和底盘膨大率, 观察茎段诱导状态。结果表明, 随着6-BA浓度的增高, 定芽诱导率呈先升高后下降趋势, 在添加3 mg?L-1 6-BA的培养基中, 定芽诱导率最高, 达84.44%; 随着6-BA浓度的增高底盘膨大率逐渐增高, 最高达88.89%, 但部分底盘裂开(表2)。综合定芽诱导率、底盘膨大率和茎段生长状况, 表明当6-BA浓度为2-3 mg?L-1时, 茎段底部产生愈伤组织较多, 部分定芽长出, 且长势较好(图1B), 为后续不定芽诱导奠定良好基础。
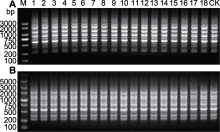
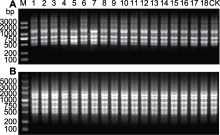
与自然环境相比, 体外培养更易引起遗传变异。因此, 在确认繁殖成功之前, 有必要对再生植株的遗传稳定性进行分析。本研究中, 我们选用RAPD和ISSR分子标记, 共扩增出161条带, 平均每个引物6.4条带(表6, 表7)。15个RAPD引物共扩增出113条清晰的条带, 长度在250-5 000 bp之间。单个RAPD引物扩增的条带数介于4-12条之间。引物S132的RAPD扩增模式如图2A所示, 引物S133的RAPD扩增模式如图2B所示。10个ISSR引物共扩增出48条清晰的条带, 长度在300-3 000 bp之间。单个ISSR引物扩增的条带数为2-6条。引物UBC854的ISSR扩增模式如图3A所示, 引物UBC880的ISSR扩增模式如图3B所示。
本文的其它图/表
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 首页
首页