|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
月季红双喜花瓣变色的化学基础及比较转录组分析
植物学报
2022, 57 (5):
649-660.
DOI: 10.11983/CBB22049
月季(Rosa hybrida)花色丰富, 是世界著名的观赏花卉。月季红双喜因其花瓣的变色特性而在市场上广受欢迎。该研究通过类黄酮和类胡萝卜素靶向代谢并结合转录组分析, 发现红双喜的黄色花瓣呈色主要源于叶绿素、类胡萝卜素以及类黄酮的积累, 红色花瓣呈色主要是花青素苷积累增加且糖苷化的结果。花青素苷合成关键基因CHI、ANS和UFGT, 以及R2R3-MYB家族的AN2-like成员在红色花瓣中的强烈表达是花青素苷积累的分子基础; 类胡萝卜素成分改变及相关基因的表达变化在红双喜花瓣变色过程中也起重要作用, 并且miRNA156可能参与其调控过程。该研究揭示了月季红双喜花瓣变色的分子和化学基础, 研究结果为观赏植物花色分子设计育种提供了重要理论依据。
表2
月季红双喜不同呈色花瓣中主要类胡萝卜素的成分含量差异
正文中引用本图/表的段落
月季(Rosa hybrida)是重要的观赏花卉, 品种多、花色丰富且适应性强, 可用于园林和切花, 也可用于盆栽或庭院栽培。花色是月季的重要观赏性状, 也是品种价值的重要体现。研究发现, 月季花瓣的呈色主要受类黄酮和类胡萝卜素的影响(温佳辛等, 2021)。类黄酮属于水溶性色素, 存在于液泡, 可决定花瓣的一系列呈色; 而类胡萝卜素属于脂溶性色素, 存在于质体, 对一些植物花瓣的橙色至黄色呈现起一定作用。类黄酮和类胡萝卜素在细胞中的合成受一系列结构基因的控制; 一些关键结构基因的表达差异是花色呈现差异化的重要分子基础(Tanaka and Ohmiya, 2008; Watkins and Pogson, 2020)。有研究表明, 类黄酮合成结构基因可在转录水平受R2R3- MYB、WD40和bHLH等转录因子的调控(Tanaka and Ohmiya, 2008); 而类胡萝卜素合成结构基因可受MADS-box、NAC、HD和AP2/ERF等转录因子的调控(Llorente et al., 2017)。此外, 类黄酮和类胡萝卜素合成的一些结构基因也会受到LncRNA和siRNA等非编码RNA的转录后调控(Bradley et al., 2017; Tu et al., 2022)。目前, 关于月季花瓣呈色的研究包括色素成分鉴定和分析, 色素合成相关结构基因的功能解析, 以及利用转基因技术对外源基因进行转化以创制具有蓝色花瓣的月季种质等方面(Katsumoto et al., 2007; 王峰等, 2017; Li et al., 2018; Sui et al., 2019; 张泰然等, 2020)。关于月季花色调控的研究, 李茂福等(2021)发现R2R3-MYB基因是月季花色形成的重要调控因子。
转录组分析在北京百迈客生物科技有限公司进行, 所用测试样本与代谢组测试一致。基于边合成边测序(sequencing by synthesis, SBS)技术对构建的cDNA文库利用Illumina HiSeq高通量测序平台获得长链非编码RNA (LncRNA)和编码RNA (mRNA)的原始数据(raw data)。将原始数据与指定的Rosa chinensis基因组(GCF_002994745.1.Rosa_chinensis. GCF_0029- 94745.1.genome.fa)进行序列比对, 并在COG、GO、KEGG、KOG、NR、Pfam、Swiss-Prot和eggNOG等数据库对编码RNA及长链非编码RNA的靶基因进行功能注释与聚类分析。对4个样本的FPKM值进行统计, 使用DEseq软件进行基因差异表达分析。检测过程中, 差异表达基因的差异倍数(fold change, FC)表示两样品(组)间表达量的比值, 采用Benjamini- Hochberg校正方法对原有假设检验得到的显著性P值(P-value)进行校正, 并且将FC≥2作为差异筛选标准。表达水平存在显著差异的基因, 称为差异表达基因(differentially expressed gene, DEG); 表达水平存在显著差异的转录本, 称为差异表达转录本(differentially expressed transcript, DET)。
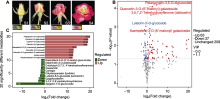
月季红双喜随着花瓣的发育, 花色逐渐由黄色变为红色。花瓣变色的趋势由外至内, 并且与萼片的开放程度有直接关系。萼片完全包住的花瓣颜色为黄色, 当萼片展开露出花瓣, 花瓣逐渐呈现红色。对不同发育阶段的红双喜花瓣色素成分在不同溶剂中的分层观察表明, 无论黄色还是红色花瓣都含有溶于下层二氯甲烷的黄绿色素成分, 为类胡萝卜素或叶绿素; 上层为溶于水的类黄酮, 并且颜色随着花瓣的开放, 逐渐由浅黄变为红色(图1A)。对不同时期同种类型呈色(黄色vs红色)花瓣进行混合, 分析其叶绿素a以及叶绿素b的含量, 显示黄色花瓣叶绿素a的含量为1.872 mg·g-1, 叶绿素b的含量为2.576 mg·g-1; 而红色花瓣叶绿素a的含量为1.698 mg·g-1, 叶绿素b的含量为2.076 mg·g-1。同时, 用靶向代谢组方法对2种类型花瓣中的类胡萝卜素和类黄酮进行测定分析。结果表明, 红双喜花瓣中含类胡萝卜素成分49种; 含量较高的α-胡萝卜素包括玉米黄质(zeaxanthin)、紫黄质(viola-xanthin)以及紫黄质-肉豆蔻酸酯-发酸酯(violaxant-hin-myristate-caprate); 含量较高的β-胡萝卜素种类主要是叶黄素(lutein)。在这些类胡萝卜素成分中, 出现差异变化的共有9种, 均是含氧类胡萝卜素类型(表2); 其中下调6种, 但是含量较低; 上调3种, 包含叶黄素和玉米黄质。红双喜花瓣中鉴定出类黄酮代谢物309种, 其中上调63种、下调37种。上调幅度较大的类黄酮物质主要为花青素苷, 包括天竺葵素-3-O-葡萄糖苷(pelargonidin-3-O-glucoside)、矢车菊素-3-O-半乳糖苷(cyanidin-3-O-galactoside)、矢车菊素-3-O-葡萄糖苷(cyanidin-3-glucoside)以及芍药素-3-O-阿拉伯糖苷(peonidin-3-O-arabinoside); 相应的下调幅度较大的主要是圣草素查尔酮(3,4,2',4',6'-pentahydroxychalcone)以及芹菜素-7-O-葡萄糖醛酸苷(apige-nin-7-O-glucuronide)等上游类黄酮代谢物(图1B)。以上分析表明, 红双喜花瓣中, 黄色花瓣贡献的色素类型包括叶绿素、类胡萝卜素和类黄酮; 而红色花瓣贡献的主要是花青素苷, 类胡萝卜素和叶绿素对红色花瓣的色调也起部分作用。
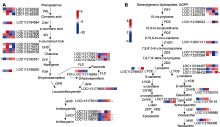
编码基因的表达差异是性状差异化的分子基础。对红双喜不同呈色花瓣编码RNA (mRNA)的表达特性进行比较分析, 其中16 857个基因无差异表达, 差异表达基因有2 250个, 上调基因1 371个、下调879个(图3A)。将差异基因进行KEGG代谢通路富集分析, 结果表明, 有3个基因参与花青素苷合成(anthocyanin biosynthesis), 14个基因参与类胡萝卜素合成(carotenoid biosynthesis), 53个基因与光信号转导过程相关(photosynthesis和photosynthesis-antenna proteins) (图3B)。
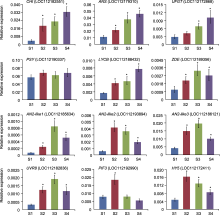
对类胡萝卜素合成结构基因的表达特性进行分析, 表明PSY、PDS以及Z-ISO等上游基因在2种呈色花瓣中无显著变化, 但是下游胡萝卜素环化酶(lycopene β-cyclase)基因LOC112188432、玉米黄质环氧化酶(zeaxanthin epoxidase)基因LOC112176116和LOC112189356, 及黄色素脱氢酶(xanthoxin dehydrogenase)基因LOC112172517等在红色花瓣中显著上调, 辣椒红素合成酶(capsorubin synthase)基因LOC1121-64457显著下调, 这些修饰基因的表达变化可能是2种类型花瓣中类胡萝卜素差异化的分子基础。
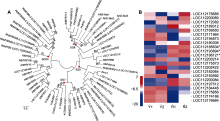
研究表明, 以R2R3-MYB为核心、MBW为单元的分子模块是植物类黄酮/花青素苷合成调控的重要方式; 与类黄酮调控相关的R2R3-MYB基因包括SG5、SG6、SG7和SG20等亚家族成员。对月季红双喜花瓣中的转录组进行分析, 共发现334个与MYB相关的转录本(Pfam数据库注释结果), 其中10个出现差异表达。将10个鉴定到的差异表达R2R3-MYB基因与月季基因组中类黄酮调控相关R2R3-MYB基因进行聚类分析, 结果表明LOC112189515、LOC112189516、LOC112172082、LOC112183574、LOC112169550和LOC112189312等属于SG5, LOC112186121、LOC112200243、LOC112193894、LOC112185634和LOC112180306五个基因成员属于SG6, LOC112- 175573和LOC112200214属于SG7, LOC112182- 064和LOC112202422属于SG20 (图5A)。对这些R2R3-MYB基因在红双喜不同呈色花瓣中的表达属性构建热图, 发现与花青素苷合成调控相关的SG6成员中有3个基因显著上调, 并将其命名为AN2-like (图5B)。
Y和R同
有研究表明, 调控植物类黄酮合成的核心元件主要是R2R3-MYB转录因子, 分布在SG5、SG6、SG7和SG20等亚家族(Zhang et al., 2019; Fu et al., 2020, 2021)。不同R2R3-MYB基因成员参与响应环境或发育信号, 并且差异化调控结构基因的表达, 从而控制类黄酮在植物不同发育时期(时间)和不同组织部位(空间), 以及成分和数量上的积累(Koes et al., 2005; Xu et al., 2015)。月季基因组中至少有6个成员聚类在SG6亚家族, 其中3个AN2-like成员(LOC 112186121、LOC112193894和LOC112185634)在红色花瓣中显著上调。但同时发现有1个聚类在SG7亚家族的成员(LOC112175573)也表达上调; 相应的, 其可能调控的下游基因FLS (LOC112170431)的表达在红色花瓣中也明显上调。因此, SG6和SG7成员可能在调控类黄酮结构基因的表达中存在竞争; 它们在细胞中的表达强弱, 以及对结构基因的调控作用的差异特性最终决定类黄酮的组成成分差异。有研究表明, miRNA156以及一些LncRNA在介导花青素苷合成中起作用(González-Villagra et al., 2017; Tu et al., 2022)。本研究对红双喜不同呈色花瓣中的差异miRNA和LncRNA也进行了分析, 其靶基因未富集到与类黄酮相关的成员, 说明红双喜红色花瓣的呈色变化调控主要受基因转录的影响, 不受非编码RNA的转录后调控。但是, 包括miRNA156在内的11条siRNA被富集到类胡萝卜素的合成过程, 说明miRNA调控可能在类胡萝卜素合成或代谢中起作用。此外, 研究2种类型花瓣中的转录组, 鉴定到多个与光信号转导相关的成员, 包括光信号受体(CRY、UVR8、phytochrome和phototropin)和光信号转导关键元件(PIF和HY5), 它们的编码基因均有表达差异。已有研究表明, 紫外光和蓝光对花青素苷的合成均有调控作用(Liu et al., 2015, 2018; Podolec and Ulm, 2018)。本研究揭示了红双喜花瓣呈色与光信号相关的线索, 但光信号参与红双喜花瓣的呈色调控尚需进一步验证。
本文的其它图/表
|
 首页
首页