
|
|
||
|
禾本科三倍体: 形成、鉴定与利用
植物学报
2021, 56 (3):
372-387.
DOI: 10.11983/CBB20166
禾本科三倍体的形成途径包括2n配子融合、倍性间杂交、多精受精和胚乳培养。其中, 2n配子融合和倍性间杂交分别为自然界和人工合成三倍体的主要途径。该文介绍了形态学观测、染色体分析、流式细胞术和分子标记等倍性鉴定方法在禾本科三倍体中的应用及其优缺点。目前, 三倍体在禾谷类作物中无直接应用价值, 但可作为通往多倍体、非整倍体和转移异源基因的遗传桥梁。多年生禾本科三倍体(特别是异源三倍体)在饲草或能源作物中已得到广泛应用, 在该类型禾本科作物中均可直接尝试三倍体育种。多倍体的三倍体育种和无融合生殖三倍体育种可作为未来禾本科三倍体的研究方向。三倍性胚乳培养可以一步合成三倍体, 多精受精可以实现遗传上3个不同基因组的一步融合, 在三倍体研究中应予以重视。鉴于2n配子融合、多精受精的稀有特性和倍性间杂交、胚乳培养频繁的染色体变异, 高通量三倍体鉴定技术的发展将是三倍体研究实现突破的关键。 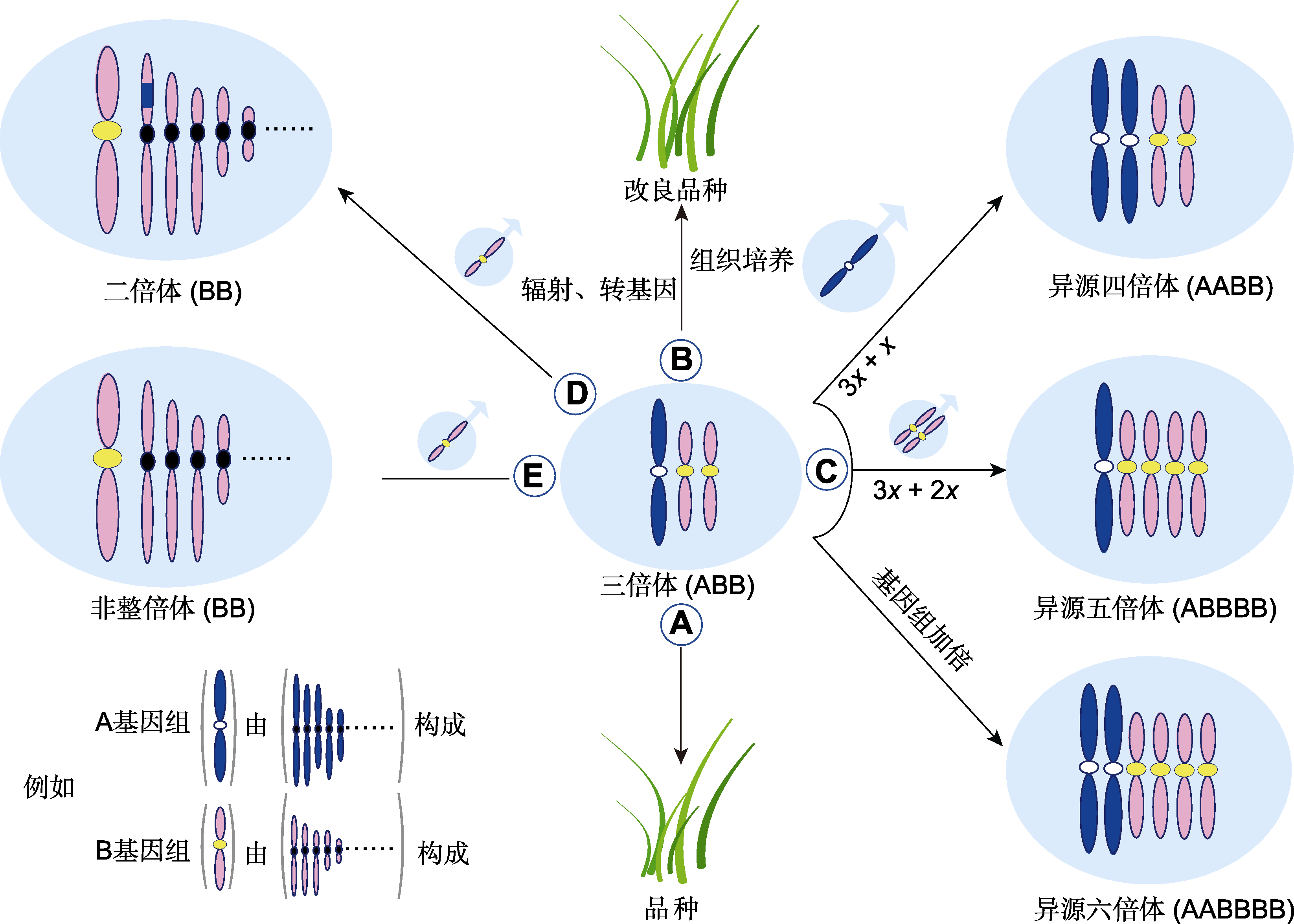 View image in article
图3
禾本科三倍体的利用
(A) 三倍体作为品种直接利用; (B) 通过辐射、转基因或组织培养等技术改良三倍体品种; (C) 作为通往更高倍性的“三倍体桥”; (D) 转移异源基因; (E) 通过3x/2x合成非整倍体
正文中引用本图/表的段落
三倍体通过保留用于生产种子的光合作用能量来促进营养生长, 通常具有更强的营养生长优势(Hoshino et al., 2011)。因此, 禾本科三倍体育种在草坪草、能源草和牧草上收获颇丰(表1; 图3A)。与二倍体相比, 三倍体的少花或无花特性有利于草坪管理和提升草坪观赏价值。在无融合生殖早熟禾(Poa pratemis)中, 筛选出源于2n配子的抗病三倍体草坪草种质(Pepin and Funk, 1971)。三倍体比二倍体海滨雀稗(Seashore paspalum, 2n=2x=20)的叶色更深(Schwartz et al., 2013)。通过四倍体普通狗牙根(2n=4x=36)与二倍体非洲狗牙根(C. transvaalensis, 2n=2x=18)杂交育成了Tifway、TifEagle和Tifgreen等知名三倍体狗牙根品种(2n=3x=27), 成为三倍体草坪草育种的杰出典范。三倍体奇岗(M. × miscanthus, 2n=3x=57)具有多年生、C4高光效、低氮需求、较耐低温和高生物量等特性, 是最重要的第二代能源植物, 现已成为能源模式植物(潘志军等, 2017)。三倍体鸭茅苗期生长速度快于双亲(钟声, 2006)。三倍体热研4号王草第3年丰产期干物质产量比母本象草 (Pennisetum purpureum)增产32.8% (刘国道等, 2012)。玉草1号由二倍体玉米与四倍体多年生大刍草杂交而成, 分蘖力强、生物量高且具再生性(任勇等, 2007)。当前生产中利用的禾本科三倍体能源草和牧草(表1)大多具有如下特性: (1) 利用异源三倍体的亚种间杂种优势; (2) 多年生高大禾草; (3) 易于无性繁殖。因此, F. apennina (2n=4x=28)与F. pratensis (2n=2x=14)合成的三倍体显示出生物量杂种优势, 但因杂交结实率低, 须提高结实率或采取无性扩繁技术, 否则难以应用(Boller and Kopeck, 2020)。高粱属和斑茅(Eriarthus arundinaceus)等高大禾草目前尚无三倍体品种, 可尝试三倍体育种。
对于有直接利用价值的三倍体, 可通过组培、诱变和转基因等技术进一步改良(图3B)。MS-Supreme源于Tifgreen的自然突变, 与Tifgreen相比其叶片和茎节节间缩小、变短(Krans et al., 1999)。在Tifgreen的自然突变中还选育出Tifdwarf和FloraDwarf等矮生品种; TifwayII、TifSport和TifEagle则由相应三倍体亲本经γ辐射诱变而成(Kamps et al., 2011)。利用体细胞无性系变异育种技术在TifEagle的愈伤组织中长时间连续继代培养能有效诱导体细胞变异(如矮化突变体); 以0.3 mol·L-1 NaCl为选择压筛选出耐盐细胞系(Lu et al., 2007)。以美洲狼尾草(Pennisetum americana)/ 四倍体象草合成的三倍体F1种子为诱变材料, 经γ辐射获得优异变异株闽牧6号(陈钟佃等, 2012)。华南1号杂交狼尾草由三倍体杂交狼尾草芽变而来(陈平等, 2004)。三倍体也可以直接作为亲本利用。在偏序钝叶草(Stenotaphrum secundatum, 2n=2x=18)的3x/ 2x杂交后代中经胚拯救选育出抗灰斑病(由稻瘟病菌(Magnaporthe grisea)引起)和耐旱草坪草新品种(Chandra et al., 2015)。奇岗天然后代群体中16个数量性状出现了丰富的变异(变异系数为14.41%- 151.85%), 在后代中筛选出3份材料, 其生物量优于奇岗(潘志军等, 2017)。通过三倍体狼尾草与摩特矮象草杂交选育出中国南方地区广泛种植的三倍体多年生牧草桂牧1号(罗宗志等, 2016)。宁牧26-2狼尾草也是由三倍体后代衍生而来(顾洪如等, 1992)。三倍体转基因品系可通过2种策略获得。(1) 直接以三倍体为受体, 导入目的基因构建转基因品系, 如三倍体狗牙根Tifeagle (Hu et al., 2005)。(2) 对于愈伤组织形成与再分化率低的三倍体, 首先需要寻找愈伤组织再分化优良的二倍体或四倍体作为转基因桥梁, 然后通过基因枪或农杆菌转化等方法导入目的基因, 最后通过倍性间杂交构建转基因三倍体, 如芒(2n=2x=38) (Wang et al., 2011)。近年来, 基因编辑技术的发展为定向改良三倍体提供了新的育种方案。
三倍体可作为通往其它倍性水平的桥梁, 即三倍体桥(triploid-bridge) (图3C)。在鸭茅三倍体后代中发现了四倍体、五倍体以及混倍体植株(钟声, 2006)。三倍体摩擦禾可产生2n雌配子, 通过3x/2x合成了四倍体(Kindiger and Dewald, 1994)。以三倍体杂交狼尾草(美洲狼尾草/象草, 2n=3x=21)的幼穗离体培养获得的胚性愈伤组织为材料, 通过秋水仙碱诱导获得六倍体(钟小仙等, 2014)。利用秋水仙碱处理象草(2n=4x=28)/珍珠粟(P. glaucum, 2n=2x=14)杂交三倍体幼苗, 体细胞加倍后形成异源六倍体(Campos et al., 2009)。用类似的方法获得M. × miscanthus六倍体(Yu et al., 2009; Ghimire et al., 2016)。水稻(AA)和高秆野生稻(O. alta, CCDD)杂交形成异源三倍体, 加倍后形成三基异源六倍体(AACCDD) (祝剑峰等, 2008)。
三倍体可促进物种间的基因交流(图3D)。例如, 水稻(AA)与4个澳洲野生稻(O. australiensis, 2n= 2x=24, EE)杂交经胚拯救获得F1, 但F1雄性不育, 其作母本未获得种子; 然而, 以四倍体水稻作母本、澳洲野生稻作父本获得三倍体(AAE), 随后用水稻作父本授粉该三倍体, 通过与三倍体回交在其后代中发现了抗褐飞虱和白叶枯病的水稻种质(Multani et al., 1994)。合成的三倍体杂交狼尾草与四倍体相比, 基因组的相对损失更少, 更有潜力作为遗传桥梁(Nunes et al., 2013)。特别地, 利用三倍体世代还可提高亲本基因组的稀释效率(党江波等, 2018)。
三倍体也是通往非整倍体的常用桥梁(图3E)。对三倍体谷子授以二倍体花粉, 在杂交后代中获得全套三体(王润奇等, 1994)。在异源三倍体水稻(AAE)/水稻的杂交后代中, BC1染色体数目变幅为28-31, BC2染色体数目变幅为24-28, 在BC2F2中获得8个附加系(2n=20Os+1Oa) (Multani et al., 1994)。在高粱(Hanna and Schertz, 1971)、粳稻(O. sativa subsp. geng) (Iwata and Omura, 1975)和籼稻(O. sativa subsp. xian) (程祝宽等, 1996)等重要禾本科作物中均有通过3x/2x构建的全套初级三体。胚乳组织常表现出有丝分裂不规则, 导致染色体桥和落后染色体进而形成非整倍体(Thomas and Chaturvedi, 2008), 如黑麦草胚乳(Norstog et al., 1969)。多花黑麦草4x/2x合成的三倍体与二倍体黑麦草杂交, 后代非整倍体占比为70.89% (56/79); 当以三倍体作父本时, 后代整倍体占比高达87.69% (741/845); 差异源于三倍体作父本时整倍体花粉有选择优势而非整倍体花粉失活(Thomas et al., 1988)。随着分子标记和测序技术的发展, 利用非整倍体进行基因定位和构建连锁图谱已无优势。在拟南芥非整倍体自交后代中的整倍体个体, 某些性状的表现与非整倍体母本保持一致(Henry et al., 2010)。Gao等(2016)证实了非整倍化在新生多倍体的基因组演化中具有重要作用, 非整倍体小麦的DNA甲基化水平和模式可以稳定地被整倍体后代继承, 这表明非整倍体可以为育种创造表观遗传变异。
本文的其它图/表
|
 首页
首页
{kind=link}