
|
|
||
|
禾本科三倍体: 形成、鉴定与利用
植物学报
2021, 56 (3):
372-387.
DOI: 10.11983/CBB20166
禾本科三倍体的形成途径包括2n配子融合、倍性间杂交、多精受精和胚乳培养。其中, 2n配子融合和倍性间杂交分别为自然界和人工合成三倍体的主要途径。该文介绍了形态学观测、染色体分析、流式细胞术和分子标记等倍性鉴定方法在禾本科三倍体中的应用及其优缺点。目前, 三倍体在禾谷类作物中无直接应用价值, 但可作为通往多倍体、非整倍体和转移异源基因的遗传桥梁。多年生禾本科三倍体(特别是异源三倍体)在饲草或能源作物中已得到广泛应用, 在该类型禾本科作物中均可直接尝试三倍体育种。多倍体的三倍体育种和无融合生殖三倍体育种可作为未来禾本科三倍体的研究方向。三倍性胚乳培养可以一步合成三倍体, 多精受精可以实现遗传上3个不同基因组的一步融合, 在三倍体研究中应予以重视。鉴于2n配子融合、多精受精的稀有特性和倍性间杂交、胚乳培养频繁的染色体变异, 高通量三倍体鉴定技术的发展将是三倍体研究实现突破的关键。 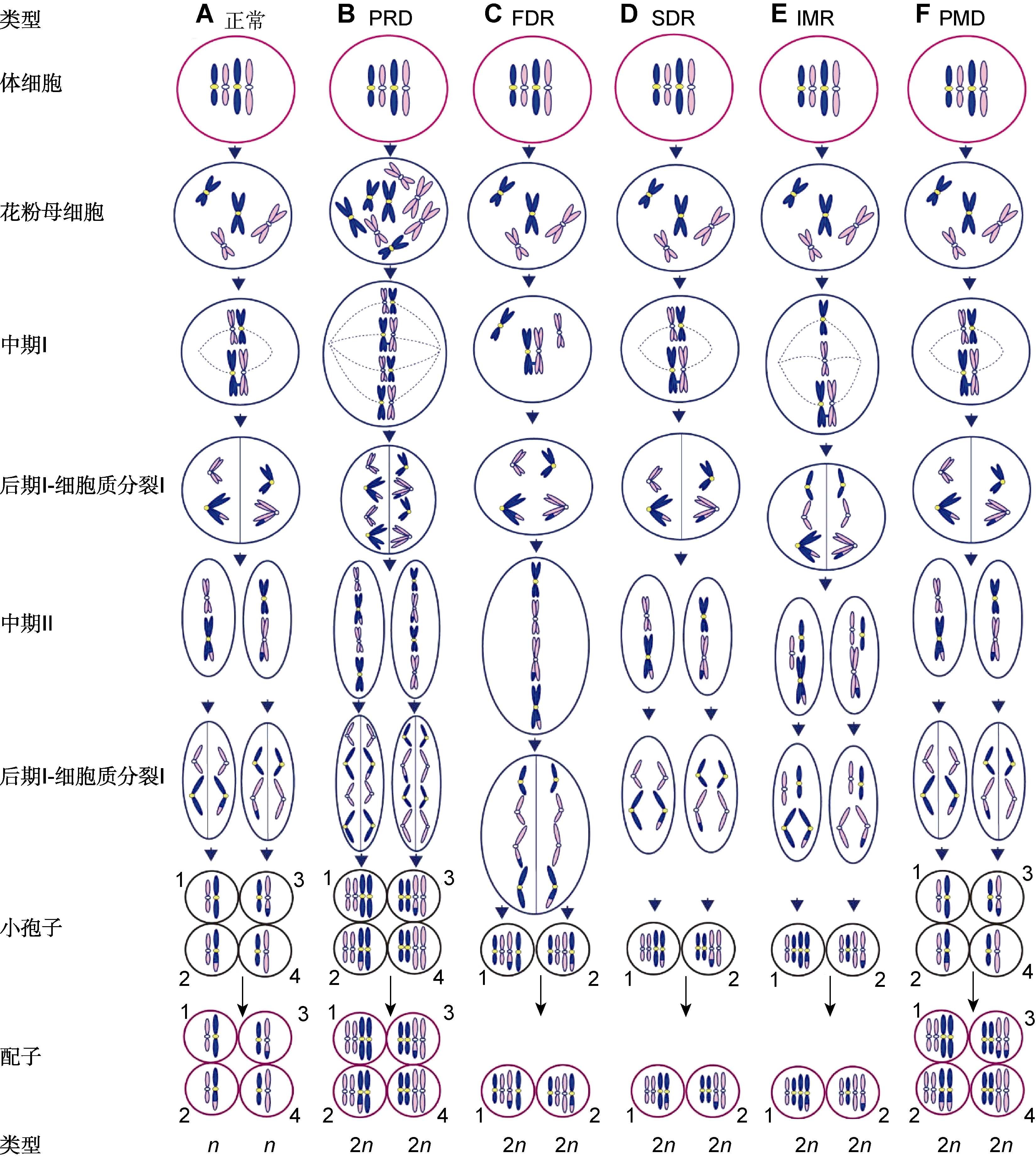 View image in article
图2
2n配子的形成机制
(A) 与体细胞相比正常配子的染色体数目减半; (B) 通过胞质融合或多核体形成的四倍体花粉母细胞产生PRD型2n配子, 如鸭茅(
正文中引用本图/表的段落
目前, 通过观测小孢子发生在植物中已发现5种2n配子形成机制(Loginova and Silkova, 2017) (图2)。减数分裂II期姐妹染色单体分离形成的FDR (first division restitution)型2n配子(图2C)在很大程度上保留了亲本的杂合性, 而减数分裂II期姐妹染色单体不分离形成的SDR (second division restitution)型2n配子(图2D)高度纯合。因此, 在转移亲本杂合性方面FDR型比SDR型更为有利(Younis et al., 2014)。Harlan和De Wet (1975)认为所有植物都以某种频率产生2n配子, 但物种间发生频率不同。二倍体水稻(Oryza sativa, 2n=2x=24)多胚苗能自发地产生三倍体(张静等, 2002)。在二倍体看麦娘(Alopecurus bulbosus, 2n=2x=14)自然群体中, 二倍体、三倍体与四倍体并存(Sieber and Murray, 1979)。在22个二倍体大麦(Hordeum vulgare, 2n=2x=14)品种中, 三倍体植株的发生率为0.01%-0.29% (Sandfaer, 1975)。在黄花茅(Anthoxanthum alpinum, 2n=2x=10)中2n配子的发生率为6.3% (Bretagnolle, 2001)。对十字花科24个物种60个群体的研究表明, 尽管2n配子先前被认为相对罕见、但普遍发生(通常个体的雄配子2n发生率<2%), 并且在特定个体中较高, 这些极少数产生高频率2n配子的个体可能在多倍体的形成中发挥关键作用(Kreiner et al., 2017)。在同一植物中, 雌雄2n配子的发生率也不一致(Ramsey and Schemske, 1998)。鸭茅(Dactylis glomerata, 2n=2x=14) 2n雌配子发生率为0.49%, 而2n雄配子发生率为0.98% (Maceira et al.,1992)。鉴于2n配子发生率低, 在2x/2x下通过单向有性多倍化(unilateral sexual polyploidization, 2x+x)形成三倍体的频率远高于通过双向有性多倍化(bilateral sexual polyploidization, 2x+2x)形成四倍体的频率。例如, 在黄花茅中2n配子形成三倍体的发生率为0.2%, 但未见四倍体发生(Bretagnolle, 2001)。
FCM检测花粉倍性为2n配子的评估提供了高通量的选择。然而, FCM难以获得高质量核荧光直方图且不能将真实的2n核(singlet)与n+n黏合细胞(doublet)区分(Kron and Husband, 2015)。2n配子的发生为稀有事件, 因此区分2n核与黏合细胞十分必要。Kron等(2015)开发并验证了使用脉冲分析(pulse analysis)对花粉核数据进行双重校正, 以便使用花粉FCM获得2n配子核的准确测量值。当在2n配子(2C DNA含量)区域中, 细胞核荧光高度和/或宽度度量显示双峰态(可指示singlets和doublets), 再结合在n配子中荧光参数的单峰分布, 可以区分2n核和黏合细胞。这些检测条件在十字花科中得到满足, 但禾本科不满足上述条件, 仍需进一步改进或开发新的方法。
卵细胞融合2个精细胞(n+n+n)形成三倍体合子( Pathways, mechanisms, and rates of polyploid formation in flowering plants 2 1998 ... 目前, 通过观测小孢子发生在植物中已发现5种2n配子形成机制(
本文的其它图/表
|
 首页
首页
{kind=link}