|
|
||||||||||||||||||||
|
禾本科作物芒遗传研究进展
植物学报
2020, 55 (5):
613-622.
DOI: 10.11983/CBB19236
芒是许多禾本科作物穗部的重要结构, 不仅可以作为区分不同品种以及基因定位的重要形态标记, 而且在禾谷类作物的种子传播、籽粒灌浆、蒸腾作用及产量形成等方面起重要作用。该文综述了小麦(Triticum aestivum)、大麦(Hordeum vulgare)和水稻(Oryza sativa)芒的结构、功能与遗传调控机制研究进展, 以期为芒性状遗传机理的进一步研究及其在育种中的应用提供参考。
表1
小麦中已定位的控制芒功能的基因
正文中引用本图/表的段落
芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能。芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势。首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(巴青松等, 2010)。其次, 芒的维管束与籽粒直接相连, 有利于光合产物运往最近的籽粒, 具有运输距离短和速度快的特性, 该特性既有利于促进光合作用, 又能促进同化物的转运(Evans and Rawson, 1970; Evans et al., 1972; Teare et al., 1972; Olugbemi et al., 1976; Olugbemi, 1978)。此外, 小麦发育后期旗叶衰老且光合速率开始下降, 芒不易老化的特性使其仍具有较高的光合速率(Teare et al., 1972; 王忠等, 1993; 李寒冰等, 2002)。在干旱条件下, 叶片的光合速率显著降低, 而芒的光合速率仍能保持相对稳定的状态, 因此干旱条件下有芒小麦相较于无芒小麦增产约1/4 (Evans et al., 1972)。此外, 芒不存在光合午休现象(杜斌等, 2010)。研究表明, 芒的光合产物约占整个穗部光合产物的1/3, 光合量占整个穗部的1/2 (李晓娟, 2006)。在大麦中, 芒作为穗部主要的光合器官, 优先表达与光合作用相关的基因(Abebe et al., 2009)。
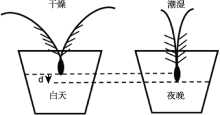
芒表面的气孔占穗部气孔数的1/2。在相同条件下, 去芒穗的蒸腾作用比未去芒穗低约75%。在炎热干旱条件下, 有芒植株通过增强蒸腾作用, 迅速降低温度, 增强吸水动力, 促进根系下扎土壤以吸收更多的水分和矿物质, 提高水分利用率, 促进水分向穗部转运, 增强植株抗旱能力(张永平等, 2006; 巴青松等, 2010; Maydup et al., 2014)。研究表明, 干旱条件下, 大麦芒中抗旱相关基因的表达量高于其内稃和外稃。因此, 在干旱和半干旱环境下, 芒对禾谷类作物籽粒产量的维持具有重要意义(Abebe et al., 2010)。
研究表明, 有芒作物的产量一般显著高于同类无芒作物, 增产约1/10 (Biscoe et al., 1973; Motzo and Giunta, 2002; Guo and Schnurbusch, 2016)。早期的研究多采用剪芒处理的方法。陈培元和李英(1981)对不同小麦品种进行剪芒处理, 发现千粒重均有下降。Rebetzke等(2016)通过构建小麦有芒、无芒近等基因系, 发现有芒小麦产量显著高于无芒小麦。大麦中的芒也通过其光合作用促进产量的增加, 芒缺失则同样伴随其产量显著下降(Milner et al., 2019)。在水稻(Oryza sativa)中, 大量与芒相关的基因不仅影响芒的发生发育, 而且对水稻穗粒数、粒长及单株分蘖等性状也有一定影响, 而这些性状都对水稻产量形成起关键作用(Luo et al., 2013)。
在普通小麦中, B1 (5AL)、B2 (6BL)和Hd (4AL)是目前已知的芒显性抑制基因, 它们的不同组合导致芒表型发生变化(Sourdille et al., 2004; Mackay et al., 2014)。3个隐性等位基因b1、b2和hd的纯合子为有芒表型, 1个显性等位基因的存在会抑制芒的伸长和发育, 含有2个显性抑制等位基因的植株是无芒表型(Antonyuk et al., 2012) (表1)。
研究表明, 小麦B1基因对芒长度的影响最强, 其突变体的表型为穗基部和中部产生短芒或无芒, 芒最长仅为1 cm (Yoshioka et al., 2017; Huang et al., 2020; Wang et al., 2020; DeWitt et al., 2020)。对B1近等基因系不同材料间的小穗数、粒长、粒重和容重进行统计, 发现无芒个体的小穗数高于有芒个体, 但有芒个体的粒重和容重较无芒个体分别增加了6.3%和11.6%, 表明B1基因与小麦小穗数、粒重和容重显著相关(DeWitt et al., 2020)。
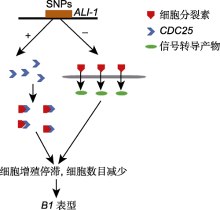
中国科学院遗传与发育生物学研究所张爱民研究团队初步将B1定位于5A染色体长臂125 kb的物理区间内, 其中包含2个基因(TraesCS5A02G542800和TraesCS5A02G542900)。表达谱和序列分析表明, 短芒材料中TraesCS5A02G542800的表达量远高于长芒材料, 而TraesCS5A02G542900在短芒和长芒材料间表达量无显著差异。测序结果表明, TraesCS5A02G542900序列在短芒和长芒材料中一致, TraesCS5A02G542800编码区序列一致, 但在启动子区有5个相同差异的SNP, B1/B1基因型材料表现为TAGA单体型, b1/b1基因型材料表现为CGAG单体型。转基因功能验证进一步表明, TraesCS5A02G542800启动子为B1芒长抑制位点的控制基因, 将其命名为ALI-1 (Wang et al., 2020)。
ALI-1编码1个C2H2锌指转录因子, 具有转录抑制活性。对B1近等基因系的芒进行纵切观察, 结果显示两者细胞长度无显著差异, 但短芒材料细胞数目仅为长芒材料的1/10, 表明短芒材料芒原基的细胞分裂被抑制。顺式调控元件预测结果表明, ALI-1区间内SNPs导致该基因缺失顺式元件BOXCPSAS1、LTRE1HVBLT49、SORLIP2AT和SITEIIATCYTC, 这些顺式元件参与调控分生组织和增殖细胞的基因表达(Hudson and Quail, 2003; Welchen and Gonzalez, 2006; DeWitt et al., 2020; Huang et al., 2020; Wang et al., 2020)。启动子中的SNPs上调了B1的表达, 导致CDC25微量表达。CDC25是细胞周期负调控因子, 可抑制细胞分裂。在植物体内, 细胞周期调节因子与植物激素相互作用。CDC25同源物被认为是细胞分裂素作用的早期靶点, 因此CDC25降低了细胞分裂素的含量(Lipavská et al., 2011; Wang et al., 2020)。在降低细胞分裂素含量的同时, ALI-1抑制细胞分裂素的信号转导, 导致细胞增殖停滞, 细胞数量减少, 进而使芒的伸长受到抑制, 表现为极显著的芒抑制作用(Wang et al., 2020) (图4)。此外, 单体型CGAG还表现出粒长和千粒重显著增加。
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成。An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(Takumi et al., 2000)。An-2基因编码的OsLOGL6蛋白通过促进野生稻细胞分裂素合成而促进芒伸长(Gu et al., 2015)。研究表明, 在水稻芒的驯化过程中, An-1基因优先被驯化, An-2基因紧随其后(Zhang et al., 2019)。
芒作为禾本科作物典型的穗部形态特征, 是不可忽视的关键性状。基于现有的研究, 探讨不同作物中芒发育相关基因之间如何进行互作, 以及不同物种中芒调控基因的作用方式及分子机制, 对于我们理解禾本科作物芒的产生和发育具有重要意义。在水稻中, An-1、An-2、LABA1、GAD1、GLA、TOB1和DL都通过促进细胞分裂或提高细胞分裂素的含量对芒发育或相关功能进行调控。在小麦中, 有证据表明B1可能通过抑制生长素相关途径而抑制芒的发育和细胞增殖。在大麦中, Lks2短芒突变通过影响生长素进而影响芒细胞的分裂。由此推测, 细胞分裂素及生长素的含量通过影响芒原基细胞分裂活性, 进而促进芒的伸长和芒刺形成, 是影响芒生长发育的主要因素。但禾本科作物特有器官——芒的产生和生长发育的分子机制还有待深入研究阐明。
研究表明, 小麦B1基因对芒长度的影响最强, 其突变体的表型为穗基部和中部产生短芒或无芒, 芒最长仅为1 cm (
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
研究表明, 小麦B1基因对芒长度的影响最强, 其突变体的表型为穗基部和中部产生短芒或无芒, 芒最长仅为1 cm (
d: Depth ... Effect of awns and drought on the supply of photosynthate and its distribution within wheat ears 2 1972 ... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
研究表明, 小麦B1基因对芒长度的影响最强, 其突变体的表型为穗基部和中部产生短芒或无芒, 芒最长仅为1 cm (
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因( An eight-parent multiparent advanced generation inter- cross population for winter-sown wheat: creation, properties, and validation 1 2014 ... 在普通小麦中, B1 (5AL)、B2 (6BL)和Hd (4AL)是目前已知的芒显性抑制基因, 它们的不同组合导致芒表型发生变化( The contribution of the awns of bread wheat (Triticum aestivum L.) to grain filling: responses to water deficit and the effects of awns on ear temperature and hydraulic conductance. 2 2014 ... 气孔的主要作用是与外界进行气体交换, 与旗叶相比, 芒的气孔总数少且体积小(
研究表明, 小麦B1基因对芒长度的影响最强, 其突变体的表型为穗基部和中部产生短芒或无芒, 芒最长仅为1 cm (
研究表明, 小麦B1基因对芒长度的影响最强, 其突变体的表型为穗基部和中部产生短芒或无芒, 芒最长仅为1 cm (
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
本文的其它图/表
|
 首页
首页