|
|
||||||||||||
|
植物基因表达调控与进化机制研究进展
植物学报
2025, 60 (4):
621-639.
DOI: 10.11983/CBB24175
功能基因表达是连接基因编码信息与蛋白质产物的一个基本生命过程, 基因表达水平被视为介于基因型与表现型之间的一种数量性状, 在植物应对气候和环境变化时发挥重要作用。该文首先系统综述了植物基因表达调控因子研究进展, 包括转录因子和小RNA等在基因表达调控中的作用。其次, 探讨了基于基因表达数据进行全基因组关联分析(GWAS)估计调控因子基因的表达数量性状基因座(eQTLs)位置以及该方法的局限性。随后从理论上分析了在突变、漂变、选择和迁移过程中的种内基因表达变异与检验方法, 在突变-漂变过程中以及在基于系统发育树的漂变-选择过程中的种间基因表达进化与检测方法。最后, 探讨了植物交配系统对基因表达进化的调控, 自交降低了有效群体大小、突变率、基因重组及外源花粉竞争, 改变了配子与合子阶段的自然选择功效等, 从而间接调控种内基因表达变异和种间基因表达进化。全文综合评述了目前的理论和实际研究进展及存在的问题, 有助于深入理解植物基因表达调控和进化机制。
表2
基于种间基因表达进化的选择检验
正文中引用本图/表的段落
长链非编码RNA是一类长度超过200 bp的非编码RNA分子, 其在转录、转录后和翻译水平等阶段调控植物基因表达。环状RNA (circRNA)属于一类特殊的lncRNA。lncRNA通过与DNA、RNA或蛋白质相互作用, 在转录水平上调控基因表达。在小麦(Triticum aestivum)中, lncRNA GARR2能够调控与赤霉素和生长素相关的差异表达基因, 从而影响小麦的株高和叶鞘长度等性状(胡卫国, 2020)。在白桦(Betula platyphylla)中, lncW20通过结合BpPOD59和BpCYP76G1基因的启动子, 上调其表达水平, 提高植物的抗氧化能力, 从而增强抗旱性(赵慧敏, 2022)。
基因表达水平可以用绝对和相对表达量来表示, 前者指在特定条件下, 基因的表达量可以被直接量化为一个具体的数值, 这种方法通常需要使用标准样品或已知浓度的样品做参考(Bustin, 2000; Eppler et al., 2005), 实验要求较高, 步骤较烦琐。后者则是通过比较目标RNA分子在不同样品或不同条件下的表达水平变化来估计其相对表达量(Fleige et al., 2006), 这种方法通常选择1个或多个内参基因作为对照, 以校正样品间的差异。衡量基因相对表达量常用的方法有2-ΔΔCT法(Livak and Schmittgen, 2001)和Pfaffl法(Pfaffl, 2001)。应用RNA-seq分析时, 同时测定多基因表达水平, 为消除基因长度和测序深度对基因表达水平的影响, 需将数据进行归一化处理(Evans et al., 2018), 常用RPKM (reads per kilobase per million)、FPKM (fragments per kilobase per million) (Mortazavi et al., 2008)以及TPM (transcripts per million)表示(Wagner et al., 2012)。
基因表达在生态与进化生物学方面有广泛应用。例如, Palma-Silva等(2024)利用基因表达评价竹芋科细穗竹芋属植物Ischnosiphon colombianus在亚马逊河岸森林系统中对2种环境的响应, 认为不同环境条件和洪涝灾害是种群遗传分化的驱动因素。Andrew等(2024)研究了澳大利亚17个相思树属(Acacia)树种响应热胁迫后的基因表达与环境关联性。基因表达测定也常用于分析特定基因在植物不同组织器官中的表达定位, 以及比较物种间或不同环境条件下的基因表达模式差异等(Wu et al., 2024), 依据统计学检验筛选表达差异显著的基因并进行注释, 但这类研究一般不探究基因表达调控因子。
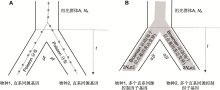
目前已有一些方法检测种间基因表达进化是否受自然选择影响(表2)。
(3) 在上述OU模型基础上, 同时考虑种内基因表达方差和种间基因表达进化方差, Rohlfs和Nielsen (2015)提出了基因表达变异和进化(expression variance and evolution, EVE)模型(表2), 该模型假设第i个物种的第k个体某个基因表达水平
(2) 目前仍以mRNA拷贝数量来度量基因表达水平, 已提出了多种基因表达的计算方法, 包括2-ΔΔCT和Pfaffl等度量方法以及数据标准化方法(RPKM、FPKM和TPM)。利用这些度量方法分析植物生态与进化问题是生物学研究的热点之一, 但是如何准确地利用基因表达水平变异定位隐含的调控因子基因仍是亟待解决的关键问题, 目前利用基因表达数据进行GWAS, 进而估计调控因子基因eQTL仍受许多因素限制, 包括样本容量、eQTL与基因组SNP连锁不平衡以及植物特定细胞或组织混合基因表达等。
(1) 类似于研究分子进化中的MK (McDonald- Kreitman) (Mcdonald and Kreitman, Overexpression of a SOC1-related gene promotes bud break in ecodormant poplars 1 2021 ... 基因表达水平类似于数量性状, 与基因本身的分子进化不同, 其表达水平的变异反映了影响该基因表达的调控因子的遗传变异(Mostafavi et al., The strength and pattern of natural selection on gene expression in rice 1 2020 ... (2) 利用两世代的调查数据进行回归分析, 测验基因表达与有关适应性数量性状的关联性来推测是否存在选择.基于适合度的自然选择变化估计选择差S, 相应的群体基因表达量的均值变化(跨越1个世代的选择响应) R, 育种学上又称为遗传增益, 根据育种者方程(breeder’s equation)估计基因表达的选择梯度参数β, 即β=R/S, 选择梯度β可用于指示表达基因的选择强度, β数值大小反映了该基因表达的选择强度.该方法在水稻育种中得到应用, 发现在湿稻田条件下, 多数基因表达处于中性或弱稳定选择(stabilizing selection), 但在干旱条件下处于强选择, 且基因表达的选择强度与cis-调控因子表达水平及网络连接性存在弱负相关(Groen et al., 2 2010 ... 式中
(1) 类似于研究分子进化中的MK (McDonald- Kreitman) (Mcdonald and Kreitman, On the probability of fixation of mutant genes in a population 1 1962 ... (1) 单个基因表达已有明确的基因类型, 表达水平受调控因子和基因本身序列进化影响, 后者在基因表达进化研究中常被忽略, 但可以从基因表达进化、选择程度(4Nes, s为突变等位基因选择系数)及基因分子进化速率( Natural selection and random genetic drift in phenotypic evolution 2 1976 ... 随着物种分化时间(t)的增长, 基因表达均值的方差就越大.实际测验时, 可应用单因素方差分析(ANOVA)估计种群间基因表达方差
(1) 类似于研究分子进化中的MK (McDonald- Kreitman) (Mcdonald and Kreitman, Ecological transcriptomics reveals stress response pathways of a ground-herb species in a waterlogging gradient of Amazonian riparian forests 1 2024 ... 基因表达在生态与进化生物学方面有广泛应用.例如, Palma-Silva等( Identification and functional annotation of long intergenic non- coding RNAs in Brassicaceae 1 2022 ... lncRNA在调控植物开花时间、器官发生以及应对生物和非生物胁迫等方面发挥作用.Shea等( 1 2019 ... 植物转录因子包括多个家族, 每个家族包含多种成员, 它们可以识别相似但略有不同的DNA序列, 从而在不同的细胞类型和生理条件下发挥不同的调控作用.以植物WRKY转录因子家族为例, 不同种类植物中WRKY基因家族成员的数量存在差异, 它们通过特异性结合启动子区域的W-box来调控基因表达.在拟南芥(Arabidopsis thaliana)中共发现90个WRKY成员, 而在沟叶结缕草(Zoysia matrella)中则发现269个WRKY成员(Tian et al., 1 2019 ... 植物转录因子包括多个家族, 每个家族包含多种成员, 它们可以识别相似但略有不同的DNA序列, 从而在不同的细胞类型和生理条件下发挥不同的调控作用.以植物WRKY转录因子家族为例, 不同种类植物中WRKY基因家族成员的数量存在差异, 它们通过特异性结合启动子区域的W-box来调控基因表达.在拟南芥(Arabidopsis thaliana)中共发现90个WRKY成员, 而在沟叶结缕草(Zoysia matrella)中则发现269个WRKY成员(Tian et al., Steroid hormones and hormone antagonists regulate the neural marker neurotrimin in uterine leiomyoma 1 2020 ... 基因表达水平类似于数量性状, 与基因本身的分子进化不同, 其表达水平的变异反映了影响该基因表达的调控因子的遗传变异(Mostafavi et al., Deep learning the cis-regulatory code for gene expression in selected model plants 1 2024 ... 总之, 基于基因表达数据进行GWAS估计调控因子仍然是一个重要途径, 是对现有转录因子进行筛选和识别分析的补充, 筛选出的调控因子通常需要进一的步实验验证(Wang et al., geiger v2.0: an expanded suite of methods for fitting macroevolutionary models to phylogenetic trees 1 2014 ... 式中 A viral suppressor modulates the plant immune response early in infection by regulating microRNA activity 1 2018 ... siRNA主要由类似于Dicer酶作用产生, 这些酶将双链RNA切割成短小的干扰RNA片段, 通过与Argonaute蛋白形成复合物(RISC)作用于目标mRNA, 导致其降解或翻译被抑制(Shamandi et al., Selection for pollen competitive ability in mixed-mating systems 1 2018 ... 式中, A new mathematical model for relative quantification in real-time RT-PCR 1 2001 ... 基因表达水平可以用绝对和相对表达量来表示, 前者指在特定条件下, 基因的表达量可以被直接量化为一个具体的数值, 这种方法通常需要使用标准样品或已知浓度的样品做参考(Bustin, Detecting signatures of selection on gene expression 3 2022 ... (1) 类似于研究分子进化中的MK (McDonald- Kreitman) (Mcdonald and Kreitman,
本文的其它图/表
|
 首页
首页