|
|
|||||||||||||||||||||||||
|
植物基因表达调控与进化机制研究进展
植物学报
2025, 60 (4):
621-639.
DOI: 10.11983/CBB24175
功能基因表达是连接基因编码信息与蛋白质产物的一个基本生命过程, 基因表达水平被视为介于基因型与表现型之间的一种数量性状, 在植物应对气候和环境变化时发挥重要作用。该文首先系统综述了植物基因表达调控因子研究进展, 包括转录因子和小RNA等在基因表达调控中的作用。其次, 探讨了基于基因表达数据进行全基因组关联分析(GWAS)估计调控因子基因的表达数量性状基因座(eQTLs)位置以及该方法的局限性。随后从理论上分析了在突变、漂变、选择和迁移过程中的种内基因表达变异与检验方法, 在突变-漂变过程中以及在基于系统发育树的漂变-选择过程中的种间基因表达进化与检测方法。最后, 探讨了植物交配系统对基因表达进化的调控, 自交降低了有效群体大小、突变率、基因重组及外源花粉竞争, 改变了配子与合子阶段的自然选择功效等, 从而间接调控种内基因表达变异和种间基因表达进化。全文综合评述了目前的理论和实际研究进展及存在的问题, 有助于深入理解植物基因表达调控和进化机制。
表1
基因表达相关调控因子与调控机制
正文中引用本图/表的段落
依据分子生物学中心法则, 基因表达调控可以发生在转录过程、转录后水平、翻译过程及翻译后水平等阶段。理论上酶蛋白和非编码RNA干扰调控都有可能作用于多个阶段。表1概括了针对不同阶段的调控因子及相关的调控机制(戴灼华和王亚馥, 2016)。下文将概述转录因子和非编码RNA对植物基因表达调控的研究进展。
转录因子是一类蛋白质分子, 在基因表达调控中的作用机制涉及多个途径, 可参与信号转导、与DNA相互作用以及在细胞特异性和发育阶段特异性表达中发挥调控作用等。许多转录因子参与细胞内外的信号转导途径, 如激素信号和环境胁迫反应(戚莹雪等, 2019; 任永娟等, 2021)。例如, 在非生物胁迫下, 通过分析HvNF-YC转录因子家族基因在耐盐型和盐敏感型大麦(Hordeum vulgare)品种中的表达量变化, 推测HvNF-YC9、10和11基因簇可能与盐胁迫响应密切相关(王菲菲等, 2023)。陈思思等(2022)对构树(Broussonetia papyrifera)转录因子BpbZIP1进行镉胁迫响应分析, 结果表明BpbZIP1基因能积极响应镉胁迫, 其表达可改善构树对镉胁迫的耐受力, 是构树响应镉胁迫的重要基因。
细胞感知胞外环境信号后, 通过信号转导途径将信息传递到胞内。转录因子接收信号并被激活, 通过结合特定的DNA序列, 招募或阻止转录复合体组装, 从而激活或抑制靶基因的转录, 上调或下调靶基因表达水平。例如, SATB1是一种通过其特殊的结构域(ULD和CUTL结构域)特异性结合DNA的转录因子(王峥, 2014)。转录因子能够感知并响应DNA的形状变化, 这在一定程度上解释了为什么某些转录因子能够识别非典型的或变异的DNA序列(Dror et al., 2014)。还有些转录因子(如FOXO1)能够以单体或二聚体的形式与DNA结合, 多聚化后增强转录因子与DNA的结合亲和力(Li et al., 2021)。通过参与信号转导并与DNA相互作用, 最终导致一些转录因子在特定的细胞类型或发育阶段特异性表达, 从而调控细胞特异性的基因表达模式(Mills and Taghert, 2012), 使不同细胞类型执行各自独特的功能。还有一些转录因子在特定的发育阶段调控植物生长发育的时序性, 涵盖从胚胎发育到器官形成, 再到生殖生长等各个重要阶段。例如, 孟令桐等(2024)对毛果杨(Populus trichocarpa) ANT转录因子进行研究, 发现PtrANT-4基因在形成层中特异性表达, 这表明ANT转录因子可能在维管形成层干细胞分裂分化过程中扮演重要角色。李佳祯等(2024)通过分析黍子(Panicum miliaceum)转录组数据, 发现转录因子PmSRS表现出组织特异性, 其在8周龄的茎、花序和根中高表达, 但在8周龄的叶片中几乎不表达。
非编码RNA (non-coding RNA, ncRNA)参与调控基因表达、细胞分化和发育等多个生物学过程。根据长度和功能的不同, 非编码RNA可分为小RNA (small RNA, sRNA)和长链非编码RNA (long non-coding RNA, lncRNA)等(陈玲玲等, 2019; Zheng et al., 2019)。这些非编码RNA分子通过与DNA、mRNA、蛋白质以及miRNA等相互作用, 在表观遗传、转录、转录后、翻译及翻译后等多个水平上调控基因表达(Zhang et al., 2019) (表1)。
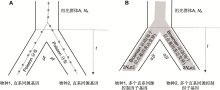
(2) 一般数量性状受多个基因表达控制, 包含细胞、组织及更高层次上的基因与基因互作效应, 基因间互作产生更复杂的网络。与此类似, 一个基因的表达水平也受到多种调控因子基因作用, 即由多基因效应组成(图1)。这些调控因子也可形成复杂的互作网络, 但更多倾向于产生细胞或组织水平上的变异。
(A) 一对直系同源基因序列进化: 在中性过程下, 物种内基因序列上的突变数用Poisson分布描述, 每个子代物种累积突变数为
(2) 利用两世代的调查数据进行回归分析, 测验基因表达与有关适应性数量性状的关联性来推测是否存在选择。基于适合度的自然选择变化估计选择差S, 相应的群体基因表达量的均值变化(跨越1个世代的选择响应) R, 育种学上又称为遗传增益, 根据育种者方程(breeder’s equation)估计基因表达的选择梯度参数β, 即β=R/S, 选择梯度β可用于指示表达基因的选择强度, β数值大小反映了该基因表达的选择强度。该方法在水稻育种中得到应用, 发现在湿稻田条件下, 多数基因表达处于中性或弱稳定选择(stabilizing selection), 但在干旱条件下处于强选择, 且基因表达的选择强度与cis-调控因子表达水平及网络连接性存在弱负相关(Groen et al., 2020)。
(1) 类似于研究分子进化中的MK (McDonald- Kreitman) (Mcdonald and Kreitman, Overexpression of a SOC1-related gene promotes bud break in ecodormant poplars 1 2021 ... 基因表达水平类似于数量性状, 与基因本身的分子进化不同, 其表达水平的变异反映了影响该基因表达的调控因子的遗传变异(Mostafavi et al., The strength and pattern of natural selection on gene expression in rice 1 2020 ... (2) 利用两世代的调查数据进行回归分析, 测验基因表达与有关适应性数量性状的关联性来推测是否存在选择.基于适合度的自然选择变化估计选择差S, 相应的群体基因表达量的均值变化(跨越1个世代的选择响应) R, 育种学上又称为遗传增益, 根据育种者方程(breeder’s equation)估计基因表达的选择梯度参数β, 即β=R/S, 选择梯度β可用于指示表达基因的选择强度, β数值大小反映了该基因表达的选择强度.该方法在水稻育种中得到应用, 发现在湿稻田条件下, 多数基因表达处于中性或弱稳定选择(stabilizing selection), 但在干旱条件下处于强选择, 且基因表达的选择强度与cis-调控因子表达水平及网络连接性存在弱负相关(Groen et al., 2 2010 ... 式中
(1) 类似于研究分子进化中的MK (McDonald- Kreitman) (Mcdonald and Kreitman, Mechanism of forkhead transcription factors binding to a novel palindromic DNA site 1 2021 ... 细胞感知胞外环境信号后, 通过信号转导途径将信息传递到胞内.转录因子接收信号并被激活, 通过结合特定的DNA序列, 招募或阻止转录复合体组装, 从而激活或抑制靶基因的转录, 上调或下调靶基因表达水平.例如, SATB1是一种通过其特殊的结构域(ULD和CUTL结构域)特异性结合DNA的转录因子(王峥, Genome-wide identification and expression analysis of the SRS gene family in Panicum miliaceum 1 2024 ... 细胞感知胞外环境信号后, 通过信号转导途径将信息传递到胞内.转录因子接收信号并被激活, 通过结合特定的DNA序列, 招募或阻止转录复合体组装, 从而激活或抑制靶基因的转录, 上调或下调靶基因表达水平.例如, SATB1是一种通过其特殊的结构域(ULD和CUTL结构域)特异性结合DNA的转录因子(王峥, 黍子SRS基因家族的全基因组鉴定与表达分析 1 2024 ... 细胞感知胞外环境信号后, 通过信号转导途径将信息传递到胞内.转录因子接收信号并被激活, 通过结合特定的DNA序列, 招募或阻止转录复合体组装, 从而激活或抑制靶基因的转录, 上调或下调靶基因表达水平.例如, SATB1是一种通过其特殊的结构域(ULD和CUTL结构域)特异性结合DNA的转录因子(王峥, On the theories of plant mating system and molecular evolution and their applications 3 2023 ... 已知交配系统演化对植物生态与进化以及物种形成等方面均有重要影响, 在植物遗传育种、资源保护和遗传多样性等研究中具有重要作用.前文讨论了4种基本动力对种内基因表达变异和种间基因表达进化的影响以及相关检测方法, 交配系统可与任一进化动力互作(Muyle and Marais,
本文的其它图/表
|
 首页
首页