|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
菊花品种万代风光再生及遗传转化体系的建立
植物学报
2025, 60 (4):
597-610.
DOI: 10.11983/CBB24151
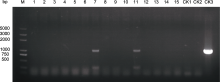
菊花品种万代风光(Chrysanthemum × morifolium ‘Wandai Fengguang’)的色素背景适宜利用分子育种技术调控花瓣铁离子浓度进而培育蓝色花, 且其在夏秋两季均可开花, 是研究菊花开花期分子调控机理的重要材料, 但缺少高效的再生体系和遗传转化体系。以该品种为实验材料, 研究不同外植体类型和植物生长调节剂组合对其再生的影响, 并探讨农杆菌介导的遗传转化方法中相关因素对遗传转化效率的影响。结果表明, 适宜菊花万代风光再生的最适外植体为茎间薄层, 最适培养基为MS+1.5 mg∙L-1 6-BA+0.6 mg∙L-1 NAA, 分化率为70.06%, 不定芽生成系数为3.37; 实验确定茎间薄层分化和不定芽生根的卡那霉素选择压分别为7.5 mg∙L-1和5.0 mg∙L-1。预培养1天、OD600=0.8、处理5分钟及黑暗条件下共培养3天为最佳遗传转化体系。经过卡那霉素筛选共获得抗性苗15株, PCR鉴定发现2株阳性苗, 转化效率为13.33%。研究结果为利用这一独特品种资源解析菊花基因功能和进行定向改良的分子育种奠定了基础, 也为其它菊花品种的再生和转化体系建立提供参考。
表1
愈伤组织诱导及不定芽分化培养基配方
正文中引用本图/表的段落
除飞燕草素途径外, 矢车菊素(cyanidin, Cy)途径也是花瓣呈现蓝色的重要机制。矢车菊(Centaurea cyanus)和大花绿绒蒿(Meconopsis grandis)均能基于矢车菊素途径产生靓丽的蓝色花, 这一过程中铁离子的参与至关重要(Takeda et al., 1996, 2005; Tanaka et al., 2001; Shiono et al., 2005)。在基于Dp途径的郁金香(Tulipa gesneriana)花瓣中瞬时表达TgVIT1基因可以提高液泡中铁离子浓度, 从而将紫色细胞转化为蓝色细胞(Shoji et al., 2007, 2010; Momonoi et al., 2009)。同样, 在基于Cy途径的蓝色矢车菊液泡铁离子转运蛋白CcVIT的氨基酸替换导致蓝色花转变为紫色(Yoshida and Negishi, 2013)。Li等(2024)发现, 菊花品种万代风光的总黄酮提取液在加入适当浓度铁离子时可变为蓝色, 其活体花瓣细胞也能够在Fe3+参与下变蓝, 证明基于Cy途径培育蓝色菊花的可行性。同时, 菊花万代风光在夏季和秋季均能开花, 开花早且花期长, 是研究菊花开花期的重要材料。因此, 为了培育蓝色菊花并实现花期的定向改良, 建立菊花品种万代风光高效且稳定的再生及遗传转化体系尤为重要。
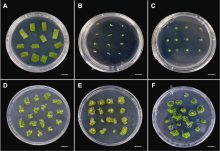
选择6-BA和激动素(kinetin, KT)为主要细胞分裂素类植物生长调节剂, 萘乙酸(naphthylacetic acid, NAA)和2,4-D为主要生长素类植物生长调节剂, 共设置4种植物生长调节剂组合, 每种组合中设置3种不同浓度配比, 共计12种培养基(表1)。每处理接种16-20个外植体, 重复3次。培养35天后统计分化率和不定芽生成系数。
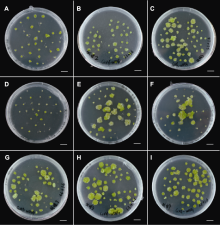
将茎间薄层分别接种于含0、2.5、5、7.5、10和15 mg?L-1卡那霉素(kanamycin, Kan)的A3培养基(表1)中。共6种Kan浓度, 每种浓度设3次重复, 每次重复接种16个茎间薄层。培养35天后统计外植体出愈率和分化率, 筛选出有效抑制外植体分化的Kan浓度。取最适分化培养基中长出的不定芽, 分别接种于含0、3、5、7.5、10和12 mg?L-1 Kan的MS培养基中。每种浓度接种4个不定芽, 重复4次。培养35天后统计不定芽的生根率, 筛选出抑制不定芽生根的最适Kan浓度。
将茎间薄层置于A3培养基(表1)上进行预培养, 预培养时间分别为2、1和0天。
侵染结束后, 用滤纸吸去多余菌液, 将外植体置于最佳再生培养基中, 分别遮光共培养1、2和3天, 用400 mg?L-1羧苄溶液冲洗10分钟, 无菌水冲洗3分钟, 用无菌滤纸吸干水分, 置于含7.5 mg?L-1 Kan和400 mg?L-1羧苄青霉素的A3培养基(表1)上。
A1-A12同
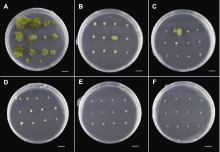
(A)-(C) 叶片、叶柄和茎间薄层在A7培养基(
(A)-(F) 茎间薄层在含0、2.5、5、7.5、10和15 mg?L-1卡那霉素的A3培养基(
本文的其它图/表
|
 首页
首页