
|
|
||
|
植物缺铁诱导型香豆素合成及其在铁吸收中的功能研究进展
植物学报
2025, 60 (3):
460-471.
DOI: 10.11983/CBB24106
香豆素类化合物是一类以苯并吡喃酮为母环结构的酚类化合物, 可分为简单香豆素和复杂香豆素, 广泛存在于自然界的高等植物中。研究表明, 缺铁条件下, 植物根部分泌的简单香豆素类化合物能够促进铁离子的吸收。该文对近年来发现和鉴定的植物缺铁诱导型香豆素合成及调控相关基因研究进展进行综述, 进一步详细阐述缺铁诱导型香豆素的生物合成、储存、分泌及其调控机制, 探讨其促进植物铁吸收的分子机制。同时, 展望该领域未来的研究方向。 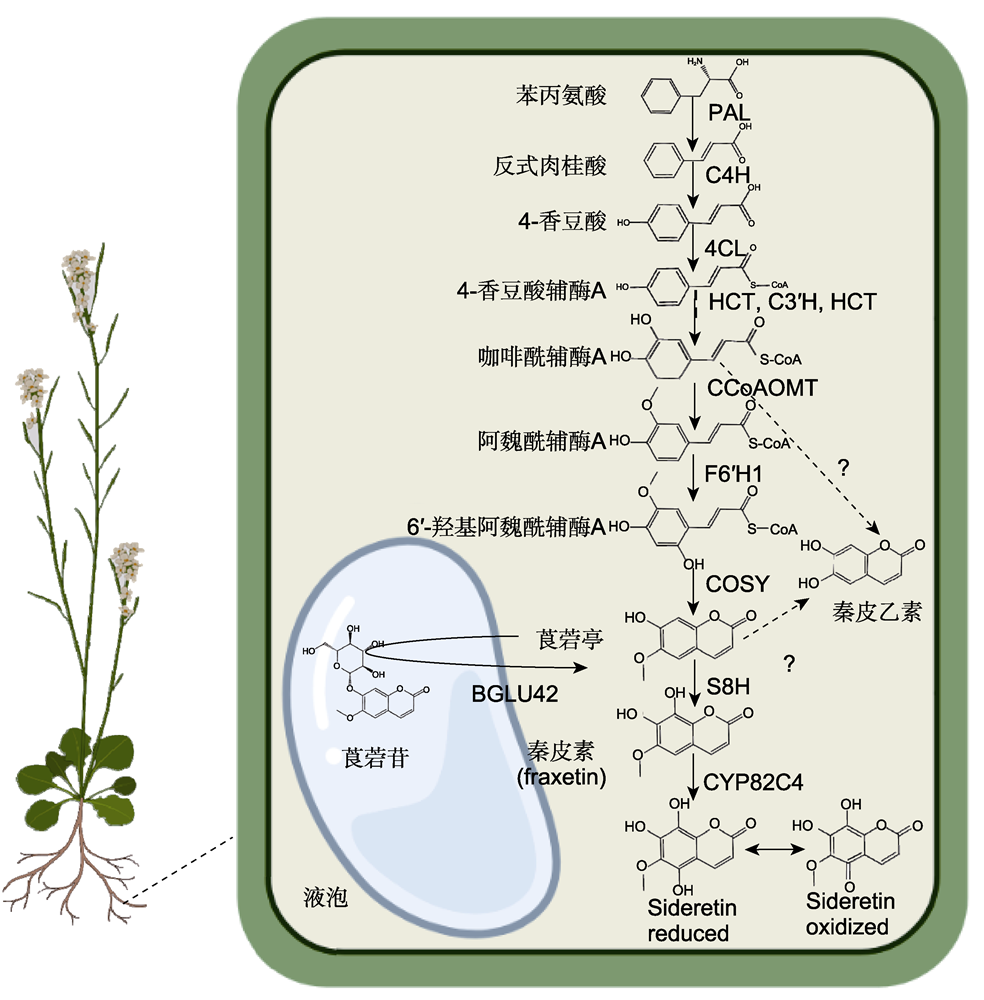 View image in article
图1
拟南芥缺铁诱导型香豆素类化合物生物合成途径
PAL: 苯丙氨酸解氨酶; C4H: 肉桂酸-4-羟化酶; 4CL: 4-香豆酸:辅酶A连接酶; HCT: 羟基肉桂酰辅酶A莽草酸/奎宁酸羟基肉桂酰转移酶; C3′H: 4-香豆酰莽草酸/奎宁酸3′-羟化酶; CCoAOMT: 咖啡酰辅酶A O-甲基转移酶; F6′H1: 阿魏酰辅酶A 6′-羟化酶; COSY: 香豆素合成酶; S8H: 莨菪亭8-羟化酶; CYP82C4: 细胞色素P450家族酶
正文中引用本图/表的段落
香豆素类化合物是一类以苯并吡喃酮为母环结构的酚类化合物, 来源于植物苯丙烷代谢途径(Waters et al., 2018)。苯丙烷代谢是最重要的植物次生代谢途径之一, 产生超过8 000种代谢物, 对植物生长发育及植物-环境互作有重要影响(Dong and Lin, 2021)。苯丙烷代谢途径中的木质素和黄酮等代谢通路已得到广泛研究, 然而直到近年, 缺铁诱导型简单香豆素的生物合成途径才得以初步解析(Robe et al., 2021b) (图1)。蛋白质组分析表明, 多个苯丙烷代谢途径关键酶在蛋白水平上响应缺铁处理(Lan et al., 2011; Pan et al., 2015)。其中包括苯丙烷代谢途径的第1个关键酶苯丙氨酸解氨酶PAL1 (phenylalanine ammonia-lyase1), 以及合成苯丙烷代谢途径的重要分支前体4-香豆酸辅酶A (4-coumaroyl-CoA)的关键酶4CL (4-coumarate:CoA ligases)。此外, 合成香豆素前体阿魏酰辅酶A (feruloyl-CoA)的关键酶羟基肉桂酰辅酶A莽草酸/奎宁酸羟基肉桂酰转移酶(hydroxycinnamoyl-CoA shikimate/quinate hydroxycinnamoyltransferase, HCT)和咖啡酰辅酶A O-甲基转移酶1(caffeoyl-CoA O-methyltransferase 1, CCoAOMT1)在蛋白水平上也响应缺铁处理(Lan et al., 2011; Pan et al., 2015) (表1)。在转录水平上, 缺铁和富铁处理之间差异基因的共表达分析表明, PAL1、PAL2、4CL1、4CL2和C4H (Cinnamate-4-hydroxylase)等香豆素前体合成相关的关键酶基因响应缺铁处理(Yang et al., 2010; Rodríguez-Celma et al., 2013; Fourcroy et al., 2014) (表1)。综上所述, 香豆素前体合成相关的关键酶基因在转录水平和蛋白水平上响应缺铁处理, 从而促进香豆素前体合成, 为进一步合成下游的香豆素类化合物奠定基础。
研究表明, 拟南芥根部可以合成并分泌具有铁活化能力的简单香豆素类化合物, 包括秦皮素fraxetin和sideretin和秦皮乙素esculetin (Rajniak et al., 2018)。首先, 香豆素前体阿魏酰辅酶A (feruloyl-CoA)在阿魏酰辅酶A 6′-羟化酶(feruloyl-CoA 6′-hydroxylase 1, F6′H1)的催化下生成6′-羟基阿魏酰辅酶A (6′-hydroxyferuloyl-CoA) (Kai et al., 2008; Schmid et al., 2014) (图1)。F6′H1是开启香豆素代谢通路的第1个关键酶, 编码F6′H1的基因功能缺失影响下游香豆素的合成。相比野生型拟南芥, f6′h1突变体在高pH或者低铁条件下生长受抑制, 呈现幼叶脉间黄化等典型的缺铁症状(Schmid et al., 2014)。进一步研究发现, 外源施加香豆素类化合物可以恢复f6′h1突变体的缺陷表型, 表明香豆素类化合物在铁吸收中具有重要作用(Schmid et al., 2014)。莨菪亭scopoletin是拟南芥中最主要的香豆素类化合物之一, 由6′-羟基阿魏酰辅酶A通过反式-顺式异构化和内酯化生成, 该反应步骤在某种程度上可以在光催化下自发进行。Vanholme等(2019)研究表明, BHAD酰基转移酶家族的香豆素合成酶(coumarin synthase, COSY)可以提高该步骤的反应效率, 并且在植物根部等无光照条件下促进莨菪亭的生物合成(Vanholme et al., 2019) (图1)。莨菪亭scopoletin是合成秦皮素fraxetin和sideretin的重要前体。莨菪亭scopoletin在莨菪亭8-羟化酶(scopoletin 8-hydroxylase, S8H)的催化下生成秦皮素(fraxetin) (Siwinska et al., 2018; Tsai et al., 2018), fraxetin进一步在细胞色素P450家族酶CYP82C4的催化下生成sideretin (图1)。然而, 关于秦皮乙素esculetin的生物合成仍有争议。Vanholme等(2019)发现香豆素合酶COSY可以在体外催化6′-hydroxycaffeoyl-CoA合成秦皮乙素esculetin, 但是该催化步骤未在植物体内得到证实。此外, Rajniak等(2018)研究发现, 在烟草(Nicotiana tabacum)中瞬时表达F6'H1基因可以诱导秦皮乙素esculetin的合成, 推测秦皮乙素esculetin可能通过莨菪亭scopoletin去甲基化合成, 其反应合成步骤有待进一步研究。COSY、S8H和CYP82C4在转录水平上受缺铁和高pH诱导(表1), 且其基因缺失突变体相比野生型表现出对缺铁和高pH更敏感, 进一步证实这些基因参与合成的下游香豆素类化合物秦皮素fraxetin和sideretin在植物响应缺铁反应中具有重要功能(Murgia et al., 2011; Rajniak et al., 2018; Siwinska et al., 2018; Tsai et al., 2018; Vanholme et al., 2019)。
PAL、C4H、4CL、HCT、C3′H、CCoAOMT1、F6′H1、COSY、S8H和CYP82C4同
本文的其它图/表
|
 首页
首页
{kind=link}