|
|
||||||||||||||||||||||||||||
|
玉米开花期调控机理及育种应用
植物学报
2024, 59 (6):
912-931.
DOI: 10.11983/CBB24127
玉米(Zea mays)是集粮食、饲料和工业原料于一身的重要农作物。开花期是作物适应不同生态环境及产量的关键决定因素。玉米开花期由营养生长时相转变和成花转变决定, 是植物内部因素(遗传因子和植物激素等)和外部环境因素共同作用的结果。鉴于玉米开花期性状的重要性, 该文从控制玉米开花期2次时相转变的组织结构基础、生理基础、遗传基础以及分子调控机理等方面系统总结了玉米开花期的遗传调控机制, 以及关键开花调控因子对玉米区域适应性的重要性, 并对玉米花期性状研究和应用的重点方向进行了讨论, 旨在加深我们对玉米开花期遗传调控的理解, 为培育广适的玉米新品种提供理论依据。
表2
玉米营养生长时相转变已知数量性状位点(QTLs)的详细信息
正文中引用本图/表的段落
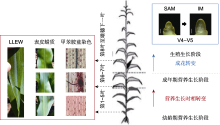
为探究调控玉米营养生长早期的遗传机制, 前人利用不同遗传背景或花期差异较大的玉米材料构建了遗传群体, 并结合分子标记图谱, 定位了一些控制营养生长时相转变的QTLs, 详细信息见表2。玉米最后一片具有蜡质的叶片是衡量玉米营养生长时相转变快慢的重要指标(Foerster et al., 2015)。基于这一表型, 进一步利用花期相近的B73与Mo17自交系构建重组自交系(recombinant inbred line, RIL)和巢式关联定位群体(nested association mapping, NAM), 通过全基因组关联分析(genome-wide association study, GWAS), 共挖掘到13个QTLs; 其中, 位于第9号染色体的SNP最为显著, 该位点对应的基因是Gl15 (Glossy15), 该基因编码AP2-like转录因子, 是zma-miR172的靶基因, 负调控营养生长时相转变(Lauter et al., 2005; Foerster et al., 2015; Xu et al., 2017); 另一个显著的SNP位于第2号染色体, 对应的基因是EDM2-like (拟南芥营养生长时相转变和成花转变关键负调控基因EDM2 (enhanced downy mildew 2)的同源基因) (Tsuchiya and Eulgem, 2010a, 2010b), 暗示EDM2-like可能参与调控玉米营养生长时相转变。此外, Xu等(2017)系统比较了长日照(long-day, LD)条件下, 玉米祖先大刍草(营养生长时相转变晚)和现代栽培玉米(营养生长时相转变早)的LLEW表型, 分别以二者典型代表8759和W22构建BC2F3群体, 再结合GWAS分析, 共鉴定到12个QTLs, 其中qVT9-1效应最大, 该位点的候选基因也对应Gl15, 其在大刍草中的表达水平远高于现代栽培玉米。进一步分析发现, Gl15的3′末端发生单核苷酸变异, 导致其在现代栽培玉米中提前终止, 表达水平降低, 从而缩短了植株的营养生长时间(Xu et al., 2017)。由此可见, 玉米营养生长时相转变由单个主效基因和多个微效基因共同调控, 其中Gl15发挥主要作用。因此, 利用营养生长时相转变差异显著的玉米种质构建群体, 结合GWAS、转录组学和代谢组学等新技术手段, 有望更快、更准确地定位到早期营养生长时相转变的关键QTLs。
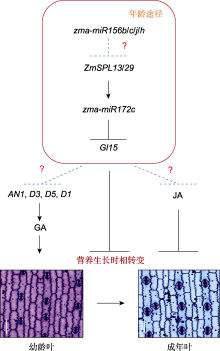
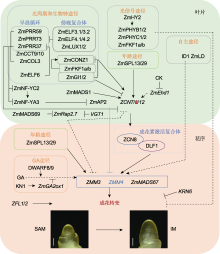
年龄途径(miR156-SPL-miR172-AP2)也是一条不依赖于光周期的开花途径(Wang et al., 2009; Wu et al., 2009)。在玉米中, zma-miR156家族成员zma-miR156b/c/g/h以及下游基因Gl15对开花起负调控作用, 而ZmSPL13和ZmSPL29能够直接促进成花转变, 从而诱导玉米开花(Lauter et al., 2005; Chuck et al., 2007; Yang et al., 2023a)。进一步分析发现, ZmSPL13和ZmSPL29蛋白通过2种机制促进玉米开花: 在叶片中能直接激活成花素基因ZCN8的表达, 以促进玉米开花; 在茎尖分生组织中能够特异性结合在花器官发育相关的MADS-box基因ZMM3/4的启动子上, 促进其表达, 从而诱导玉米成花转变(Yang et al., 2023a)。AP2转录因子ZmRap2.7和ZmAP2是玉米花期的抑制基因, 二者突变后, 植株均表现为早花(Liang et al., 2019; Su et al., 2021)。VGT1是ZmRap2.7上游的调控元件, 通过抑制ZmRap2.7的表达促进开花(Salvi et al., 2007)。此外, AG (AGAMOUS)类MADS-box基因ZmMADS69作用于ZmRap2.7上游, 抑制其表达, 从而解除ZmRap2.7蛋白对ZCN8的转录抑制, 进一步促进玉米开花(Liang et al., 2019)。
除上述5条开花途径外, 玉米花期还受到细胞分裂素(cytokinin, CK)的影响。研究发现, 外施细胞分裂素活性物质BAP时, 叶片中ZmEhd1 (Early heading date1)和ZCN8的表达被抑制, 导致植株开花延迟(Cho et al., 2022), 这表明CK在抑制玉米开花中起重要作用。此外, ZFL1和ZFL2 (拟南芥LFY (LEAFY)的同源基因)是重要的玉米花器官发育调控因子, 同时也是关键的开花整合因子。ZFL1/2的单突变体均表现为晚花, 双突变体则表现出更晚花且雄穗雌化的表型, 表明ZFL1和ZFL2功能冗余且发生功能分化(Bomblies et al., 2003)。进一步分析表明, ZFL2主要负责控制花序构型, 而ZFL1负责控制开花期(Bomblies et al., 2003)。此外, 调控玉米穗行数关键基因KNR6 (KERNEL NUMBER PER ROW6)被沉默后, 植株表现为早花; 而将其过量表达后, 则导致植株晚花(Li et al., 2023b), 这表明KRN6能够负调控玉米花期。
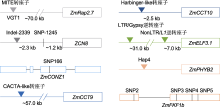
目前, 已有大量研究基于玉米花期及光周期敏感性进行的QTLs分析。研究显示, 玉米基因组上至少含有上千个SNPs和近百个基因组区域与花期性状密切相关, 这些位点几乎遍布于玉米的10条染色体上。这些QTLs能被统计方法检测出来, 预示着其背后必然有功能性的自然变异发挥作用, 其中一些QTLs在不同遗传群体或多项研究中被反复鉴定, 如ZmCCT10和ZCN8 (Hung et al., 2012; Yang et al., 2013; Guo et al., 2018; Zhong et al., 2021), 表明这些QTLs的功能变异在自然界中可能以相对较高的频率存在。然而, 仍有相当一部分QTLs只能在特定遗传群体, 尤其是在双亲连锁群体中被特异性检测到, 这暗示可能存在稀有功能变异对玉米花期起到调控作用, 那么这些QTLs的功能变异可能需要通过复杂的正向遗传学方法进行克隆和鉴定。即使以上2种情况相加, 目前真正被克隆并鉴定出功能性自然变异的花期基因仍十分有限。此外, 玉米的演化经历了漫长且复杂的育种过程, 研究人员对这些过程中的基因组选择区域进行了深入研究(Hufford et al., 2012; Liu et al., 2015a, 2015b; Wang et al., 2020; Chen et al., 2022b)。尽管不同演化过程的遗传选择基础有所不同, 但无一例外地都包含了大量花期相关基因。这些花期基因被选择下来, 背后必然存在导致基因功能改变的功能性自然变异, 这些功能变异才是玉米育种过程中真正的靶标。然而, 目前真正被克隆并鉴定的受选择功能性自然变异的基因仅有VGT1、ZmMADS69、ZCN8、ZmCONZ1、DLF1、ZmMADS67、ZmCCT9、ZmCCT10、ZmELF3.1、ZmPHYB2和ZmFKF1b (Salvi et al., 2007; Yang et al., 2013; Castelletti et al., 2014; Huang et al., 2018; Guo et al., 2018; Liang et al., 2019; Sun et al., 2020; Wu et al., 2023; Zhao et al., 2023)。此外, ZmPRR37a被认为在玉米演化过程中发挥重要作用(Yang et al., 2023a), 尽管有较强的群体遗传学证据支撑, 然而其功能性自然变异/单倍型并未鉴定清楚, 一定程度上限制了对玉米演化过程的理解及这些基因在玉米花期改良中的应用。总体而言, 鉴于玉米花期遗传调控基础的复杂性, 目前对花期基因功能性自然变异的挖掘还相对滞后。因此, 深入挖掘玉米花期调控基因的功能性自然变异将是未来玉米花期研究的重点方向之一。
本文的其它图/表
|
 首页
首页