|
|
||||||||||||||||||||||||
|
玉米开花期调控机理及育种应用
植物学报
2024, 59 (6):
912-931.
DOI: 10.11983/CBB24127
玉米(Zea mays)是集粮食、饲料和工业原料于一身的重要农作物。开花期是作物适应不同生态环境及产量的关键决定因素。玉米开花期由营养生长时相转变和成花转变决定, 是植物内部因素(遗传因子和植物激素等)和外部环境因素共同作用的结果。鉴于玉米开花期性状的重要性, 该文从控制玉米开花期2次时相转变的组织结构基础、生理基础、遗传基础以及分子调控机理等方面系统总结了玉米开花期的遗传调控机制, 以及关键开花调控因子对玉米区域适应性的重要性, 并对玉米花期性状研究和应用的重点方向进行了讨论, 旨在加深我们对玉米开花期遗传调控的理解, 为培育广适的玉米新品种提供理论依据。
表1
玉米营养生长时相转变的特性
正文中引用本图/表的段落
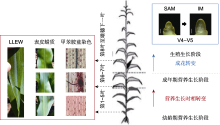
营养生长时相转变是玉米从不具备开花潜力的幼龄期向具有开花潜力的成年期的转变, 是玉米具备响应开花信号的关键转折点(Poethig, 2010)。在这一转变过程中, 玉米通常会出现多种形态结构上的变化, 包括叶片形状、叶片蜡质、表皮毛、叶片泡状细胞和中脉的形成以及角质层厚度等(Poethig, 2013) (表1)。一般而言, 成熟玉米植株的第1-5片叶为幼龄叶, 第6-7叶为转变叶, 第8叶至雄穗以下的叶片为成年叶。玉米幼龄叶表现为叶片狭长、有角质层蜡、无表皮毛、无泡状细胞带和中脉、角质层厚度小于1 μm; 经甲苯胺蓝(toluidine blue O, TBO)染色的表皮细胞呈红紫色, 细胞壁木质化程度低, 外周呈波浪状。相反, 成年叶较宽、不具有角质层蜡、上表皮着生表皮毛、叶片具有泡状细胞带和中脉、角质层厚度约为3 μm; 经TBO染色的表皮细胞呈蓝绿色, 细胞壁高度木质化, 外周锯齿状突起显著。转变叶则是从叶基部开始获得成年叶特征, 并向中部延伸, 而叶尖仍然保持幼龄叶的特征(Moose and Sisco, 1994)。此外, 最后一片具有蜡质的叶片(last leaf with epicuticular wax, LLEW)也能直接反映玉米营养生长时相转变的快慢(Foerster et al., 2015) (图2)。
光信号途径是植物感受外界环境变化进而调控植物开花的关键途径。光敏色素是植物感受红光(red light, R)和远红光(far-red light, FR)的主要光受体, 对于植物的生长发育和生理功能具有重要调节作用(Mcnellis and Deng, 1995; Franklin et al., 2003)。早期研究发现, 玉米中ELM1 (Elongated Mesocotyl1, ZmHY2, 编码线性四吡咯环化合物合成酶)突变后, 植株体内缺乏功能性光敏色素, 导致玉米在长日照条件下显著提早开花(Sawers et al., 2002)。玉米中共有6个光敏色素基因, 分别是ZmPHYA1、ZmPHYA2、ZmPHYB1、ZmPHYB2、ZmPHYC1和ZmPHYC2 (Sawers et al., 2005)。其中Zmphyb1/Zmphyb2双突变体在长日照(LD)和短日照(short-day, SD)条件下均表现出较野生型(wild type, WT)植株显著早花; 而Zmphyc1/ Zmphyc2双突变体仅在LD条件下表现出较WT植株适度早花(Franklin et al., 2003; Sheehan et al., 2007; Li et al., 2020)。此外, ZmPHYB1和ZmPHYB2在开花期的调控方面存在一定的功能分化, 其中ZmPHYB2启动子上的MITE序列能显著抑制其表达, 从而促进玉米开花(Zhao et al., 2014)。据报道, 将拟南芥phyB的104位和361位点的酪氨酸[Y]突变为苯丙氨酸[F]后, 能增强其对红光的敏感度, 从而提高phyB的活性, 并减弱拟南芥对避荫反应的响应能力(Zhang et al., 2013)。基于这一发现, 本实验室前期创制了功能增强玉米ZmPHYB1超敏材料(ZmPHYB1Y98F (对应拟南芥Y104F突变)以及ZmPHYB1Y359F (对应拟南芥Y361F突变)), 并对这些材料进行了表型调查, 结果发现幼苗期ZmPHYB1超敏材料对模拟遮阴处理(EOD-FR)的响应能力减弱, 且成熟植株的株高和穗位高均降低(Zhao et al., 2022)。
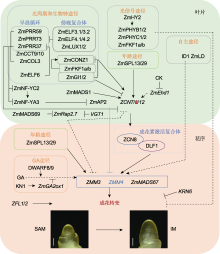
光周期和生物钟途径是调控玉米花期和光周期敏感性的核心途径。生物钟系统主要由输入途径、中央振荡器和输出途径3部分组成, 但目前玉米生物钟仅有少部分组分被鉴定出来。其中, ZmPRR37、ZmPRR59和ZmPRR73共同组成了早晨循环(morning loop); 这些基因在转录水平上表现出明显的昼夜节律, 早晨高表达, 傍晚低表达。将这3个基因突变后, 玉米植株在LD和SD条件下均出现提早开花(Yang et al., 2013, 2023b)。以上结果表明早晨循环抑制玉米开花。此外, ZmCCT9和ZmCCT10是响应光周期的开花抑制因子, 后者是水稻Ghd7的同源基因, 二者的表达模式与ZmPRR37/59/73十分相似, 均呈现早晨高表达, 傍晚低表达的昼夜节律。将ZmCCT9和ZmCCT10突变后, 玉米植株在LD条件下也提早开花(Huang et al., 2018)。本实验室前期研究发现, ZmELF3.1/3.2 (拟南芥ELF3 (EARLY FLOWERING3)的同源基因)、ZmELF4.1/4.2和ZmLUX1/2 (拟南芥LUX的同源基因)共同组成玉米的傍晚复合体(evening complex, EC), 这些基因在傍晚高表达, 早晨低表达, 具有明显的昼夜节律。将这些基因敲除后, 玉米植株在LD和SD条件下均表现为晚花。进一步分析发现, EC复合体中ZmLUX1能直接抑制下游开花抑制基因ZmPRR37、ZmPRR73、ZmCCT9、ZmCCT10和ZmCOL3的表达, 从而解除其对成花素基因ZCN7/8/12的抑制作用, 促进玉米开花(Zhao et al., 2023)。这些结果表明, EC复合体在促进玉米开花方面具有重要作用。另外, ZCN7/8/12是响应光周期的开花促进因子(Danilevskaya et al., 2008; Lazakis et al., 2011; Meng et al., 2011)。其中, ZCN8启动子区域的单核苷酸变异(SNP-1245)能直接影响成花激活因子ZmMADS1编码基因(拟南芥成花整合子编码基因SOC1 (SUPPRESSOR OF OVEREXPRESSION OF CO- NSTANS1)的同源基因)对其启动子的调控作用, 进而调控玉米开花(Guo et al., 2018)。此外, ZmGI1和ZmGI2作为生物钟中央振荡器的核心组分, 其蛋白能直接抑制ZCN8和CONSTANS-like基因ZmCONZ1的表达水平, 进而负调控玉米开花(Bendix et al., 2013; Li et al., 2023c)。而与生物钟相关的蓝光受体F-box蛋白ZmFKF1a/b (拟南芥FKF1 (FLAVINBINDING, KELCH REPEAT, F-BOX1)的同源蛋白)能够与ZmCONZ1和ZmGI1蛋白形成多聚蛋白复合体, 进而促进ZmCONZ1和ZCN8的表达, 诱导玉米开花(Chen et al., 2024)。
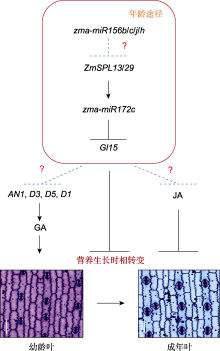
自主途径是一条不依赖于光周期的开花途径, 其中ID1 (INDETERMINATE1)是主要调控基因, 该基因编码植物特有的C2H2锌指蛋白, 主要在未成熟叶片中表达, 且其表达水平不受光周期影响(Colasanti et al., 1998; Colasanti and Sundaresan, 2000)。id1功能缺失型突变体表现叶片数增多、营养生长期延长以及开花期延迟的表型(Colasanti et al., 1998)。这表明ID1作为源自叶片的长距离成花信号, 具有促进玉米开花的作用。此外, ZmLD (Zea mays LUMINIDEPENDENS)是自主开花途径的另一个关键调控基因。ZmLD与拟南芥LD高度同源, 主要在茎尖分生组织和未成熟的花序中高表达, 将该基因在拟南芥中异位过量表达会使植株花序发育异常(van Nocke et al., 2000)。然而, 相较于其它开花调控途径, 已鉴定的自主途径调控基因还比较少, 因此仍需加强对该途径中关键组分的挖掘。
除上述5条开花途径外, 玉米花期还受到细胞分裂素(cytokinin, CK)的影响。研究发现, 外施细胞分裂素活性物质BAP时, 叶片中ZmEhd1 (Early heading date1)和ZCN8的表达被抑制, 导致植株开花延迟(Cho et al., 2022), 这表明CK在抑制玉米开花中起重要作用。此外, ZFL1和ZFL2 (拟南芥LFY (LEAFY)的同源基因)是重要的玉米花器官发育调控因子, 同时也是关键的开花整合因子。ZFL1/2的单突变体均表现为晚花, 双突变体则表现出更晚花且雄穗雌化的表型, 表明ZFL1和ZFL2功能冗余且发生功能分化(Bomblies et al., 2003)。进一步分析表明, ZFL2主要负责控制花序构型, 而ZFL1负责控制开花期(Bomblies et al., 2003)。此外, 调控玉米穗行数关键基因KNR6 (KERNEL NUMBER PER ROW6)被沉默后, 植株表现为早花; 而将其过量表达后, 则导致植株晚花(Li et al., 2023b), 这表明KRN6能够负调控玉米花期。
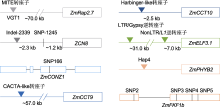
ZCN8是玉米响应光周期的成花激活子, 其启动子上的SNP-1245与开花时间高度相关。该SNP有A和G两种基因型。其中, A基因型(SNP-1245A)属于早花基因型, 携带该基因型的玉米品系中, ZCN8的表达水平较高, 表现为早花; G基因型(SNP-1245G)则相反(Guo et al., 2018)。此外, ZCN8启动子区的Indel-2339缺失同样导致玉米提早开花, 该基因型在早花温带玉米中的等位频率远高于晚花热带玉米(Guo et al., 2018)。这些结果表明, ZCN8启动子区域的SNP-1245和Indel-2339在玉米从热带地区向温带地区扩张的过程中发挥重要作用。ZmCONZ1是ZCN8的正调控基因, 其编码区第166个氨基酸与开花期密切相关, 该位点存在A和G两种基因型。其中, A基因型(SNP-166A)属于早花优异基因型, 携带该基因型的玉米品系中, ZmCONZ1表达水平高于G基因型(SNP-166G)的品系; SNP-166A基因型在温带玉米中的分布频率(58%)远高于热带玉米(4%), 特别是在高纬度北美玉米品种中的分布较多, 这表明ZmCONZ1的SNP-166A基因型在玉米适应北方高纬度地区中起促进作用(Wu et al., 2023)。此外, DLF1作为ZCN8的互作蛋白, 两者共同形成玉米FAC, 进一步促进DLF1激活下游花发育基因ZmMADS4 (ZMM4)和ZmMADS67的表达, 诱导玉米开花。研究显示, 在大刍草向现代栽培玉米的演化过程中, DLF1的3′调控区和ZmMADS67明显受到了选择(Sun et al., 2020), 这表明ZCN8/DLF1模块在促进玉米区域适应性中起关键作用。
此外, 光受体介导的光信号途径也在玉米地域适应性中发挥关键作用。例如, 红光受体基因ZmPHYB2的早花优异基因型为单倍型Hap4, 该单倍型的等位基因频率在我国玉米育种进程中随着育种年代的推进逐步提高, 而其表达水平逐渐降低(Li et al., 2023a)。蓝光受体基因ZmFKF1b在我国近现代玉米选育过程中随着育种年代的推进核酸多态性显著降低, 进一步分析表明, ZmFKF1b的早花单倍型是ZmFKF1bHap_C7-2, 携带该单倍型的玉米种质中, ZmFKF1b的表达水平较携带ZmFKF1bHap_Z58单倍型的种质高, 且开花时间提前。这主要是因为ZmFKF1bHap_C7-2单倍型玉米种质对光周期的敏感度显著降低, 使得玉米能够适应低纬度温带地区(Chen et al., 2024)。
然而, 在实际生产中, 北方长日照条件下的大部分热带种质会出现开花延迟甚至不开花的现象, 这极大地限制了热带种质在温带地区的应用。当前, 全球主要的玉米产区, 如我国的黄淮海夏玉米区、北方春玉米区和美国的黄金玉米带, 均集中在中纬度的温带地区。这是玉米从热带向温带扩张选择的结果, 也预示着对花期及光周期敏感性的选择可以实现热带种质向温带的适应性改良。研究表明, 这一过程至少涉及对VGT1、ZmMADS69、ZCN8、ZmCONZ1、DLF1、ZmMADS67、ZmCCT9、ZmCCT10、ZmELF3.1、ZmPHYB2和ZmFKF1b等重要花期基因功能变异位点的人工选择(Salvi et al., 2007; Yang et al., 2013; Castelletti et al., 2014; Huang et al., 2018; Guo et al., 2018; Liang et al., 2019; Sun et al., 2020; Wu et al., 2023; Zhao et al., 2023)。未来, 基于花期性状的QTLs、关键基因和功能性分子标记, 结合对复杂性状的全基因组选择技术、主效基因的分子标记辅助选择技术及基因编辑技术等分子育种方法, 促进玉米近源热带资源的高效改良应用, 将成为加速温带玉米种质突破的有效途径。
DPS: 散粉期; DTA: 开花期; LB: 穗下叶片数; DTS: 吐丝期; LN: 总叶片数.字体加粗表明该QTL已得到遗传验证. ... The circadian clock-associated gene gigantea1 affects maize developmental transitions 1 2013 ... 光周期和生物钟途径是调控玉米花期和光周期敏感性的核心途径.生物钟系统主要由输入途径、中央振荡器和输出途径3部分组成, 但目前玉米生物钟仅有少部分组分被鉴定出来.其中, ZmPRR37、ZmPRR59和ZmPRR73共同组成了早晨循环(morning loop); 这些基因在转录水平上表现出明显的昼夜节律, 早晨高表达, 傍晚低表达.将这3个基因突变后, 玉米植株在LD和SD条件下均出现提早开花(Yang et al., The juvenile phase of maize sees upregulation of stress-response genes and is extended by exogenous jasmonic acid 4 2016 ... 植物激素是一类由植物自身代谢产生, 在很低浓度下就能产生明显生理效应的有机信号分子(黎家和李传友,
LLEW: 最后一片具有蜡质的叶片.字体加粗表明该QTL已得到遗传验证. ... 1 2021 ... 玉米(Zea mays)是全球种植范围最广且总产量最高的谷类作物, 约占全球谷物总产量的40% (FAO, SnapShot: control of flowering in Arabidopsis 1 2010 ... 开花期是玉米适应不同生态环境种植的关键决定因素.多项研究表明, 玉米开花期是由众多微效基因共同控制的数量性状, 花期的长短往往受到遗传物质及外界环境的共同影响(Buckler et al., Phytochromes B, D, and E act redundantly to control multiple physiological responses in Arabidopsis 2 2003 ... 光信号途径是植物感受外界环境变化进而调控植物开花的关键途径.光敏色素是植物感受红光(red light, R)和远红光(far-red light, FR)的主要光受体, 对于植物的生长发育和生理功能具有重要调节作用(Mcnellis and Deng,
DPS: 散粉期; DTA: 开花期; LB: 穗下叶片数; DTS: 吐丝期; LN: 总叶片数.字体加粗表明该QTL已得到遗传验证. ... ZmGI2 regulates flowering time through multiple flower development pathways in maize 3 2023c ... 光周期和生物钟途径是调控玉米花期和光周期敏感性的核心途径.生物钟系统主要由输入途径、中央振荡器和输出途径3部分组成, 但目前玉米生物钟仅有少部分组分被鉴定出来.其中, ZmPRR37、ZmPRR59和ZmPRR73共同组成了早晨循环(morning loop); 这些基因在转录水平上表现出明显的昼夜节律, 早晨高表达, 傍晚低表达.将这3个基因突变后, 玉米植株在LD和SD条件下均出现提早开花(Yang et al.,
本文的其它图/表
|
 首页
首页