
|
|
||
|
玉米开花期调控机理及育种应用
植物学报
2024, 59 (6):
912-931.
DOI: 10.11983/CBB24127
玉米(Zea mays)是集粮食、饲料和工业原料于一身的重要农作物。开花期是作物适应不同生态环境及产量的关键决定因素。玉米开花期由营养生长时相转变和成花转变决定, 是植物内部因素(遗传因子和植物激素等)和外部环境因素共同作用的结果。鉴于玉米开花期性状的重要性, 该文从控制玉米开花期2次时相转变的组织结构基础、生理基础、遗传基础以及分子调控机理等方面系统总结了玉米开花期的遗传调控机制, 以及关键开花调控因子对玉米区域适应性的重要性, 并对玉米花期性状研究和应用的重点方向进行了讨论, 旨在加深我们对玉米开花期遗传调控的理解, 为培育广适的玉米新品种提供理论依据。 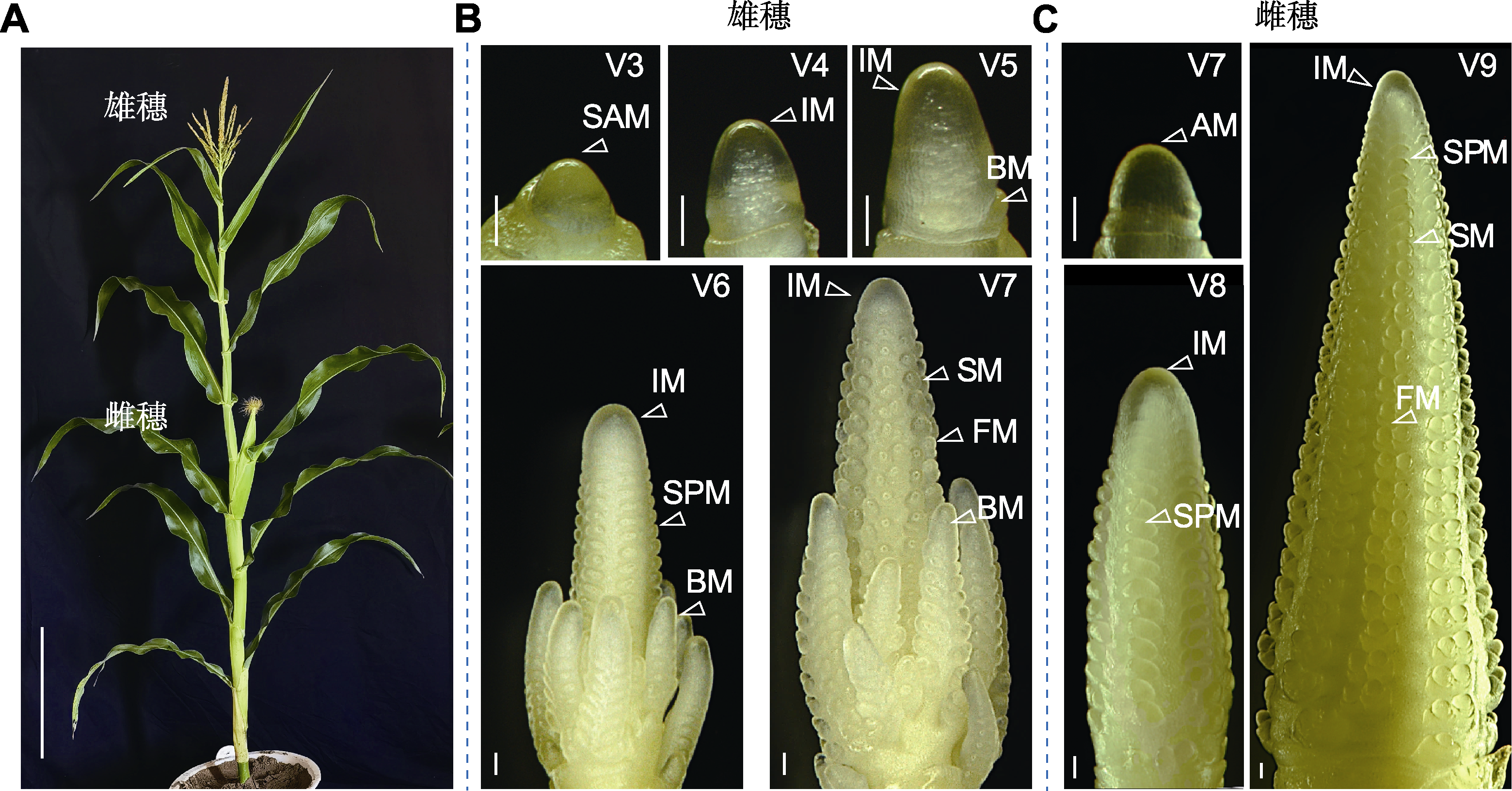 View image in article
图2
玉米雌雄花序动态发育示意图
(A) 玉米(自交系B73)为雌雄同株异花的植株(bar=30 cm); (B) 玉米雄花序发育进程(bars=100 μm); (C) 玉米雌花序发育进程(bars=100 μm); SAM: 茎尖分生组织; IM: 花序分生组织; BM: 分枝分生组织; SPM: 小穗对分生组织; SM: 小穗分生组织; FM: 小花分生组织; AM: 叶腋分生组织。V3-V9: 3至9叶期。
正文中引用本图/表的段落
营养生长时相转变是玉米从不具备开花潜力的幼龄期向具有开花潜力的成年期的转变, 是玉米具备响应开花信号的关键转折点(Poethig, 2010)。在这一转变过程中, 玉米通常会出现多种形态结构上的变化, 包括叶片形状、叶片蜡质、表皮毛、叶片泡状细胞和中脉的形成以及角质层厚度等(Poethig, 2013) (表1)。一般而言, 成熟玉米植株的第1-5片叶为幼龄叶, 第6-7叶为转变叶, 第8叶至雄穗以下的叶片为成年叶。玉米幼龄叶表现为叶片狭长、有角质层蜡、无表皮毛、无泡状细胞带和中脉、角质层厚度小于1 μm; 经甲苯胺蓝(toluidine blue O, TBO)染色的表皮细胞呈红紫色, 细胞壁木质化程度低, 外周呈波浪状。相反, 成年叶较宽、不具有角质层蜡、上表皮着生表皮毛、叶片具有泡状细胞带和中脉、角质层厚度约为3 μm; 经TBO染色的表皮细胞呈蓝绿色, 细胞壁高度木质化, 外周锯齿状突起显著。转变叶则是从叶基部开始获得成年叶特征, 并向中部延伸, 而叶尖仍然保持幼龄叶的特征(Moose and Sisco, 1994)。此外, 最后一片具有蜡质的叶片(last leaf with epicuticular wax, LLEW)也能直接反映玉米营养生长时相转变的快慢(Foerster et al., 2015) (图2)。
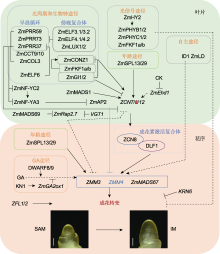
成花转变是玉米植株从营养生长向生殖生长的转变, 即茎尖分生组织(shoot apical meristem, SAM)转变为花序分生组织(inflorescence meristem, IM) (图1), 标志着玉米具备了开花能力(B?urle and Dean, 2006; Poethig, 2010)。SAM由多功能干细胞群构成, 负责产生茎、叶和花等器官。当茎尖分生组织处于SAM阶段时, 植株处于营养生长阶段; 当SAM转变为IM时, 玉米花器官发育正式起始, 植株进入生殖生长阶段。玉米是一种雌雄同株异花授粉作物, 成熟玉米植株的雄穗位于植株顶部, 而雌穗位于植株中部叶腋处(图2A)。玉米的雌雄穗具有相似的发育模式, 而IM是形成雌雄穗的前体, 也是雌雄穗命运决定的关键。当雄穗形成时, IM转变为分枝分生组织(branch meristem, BM), 随后分化出小穗对分生组织(spikelet-paired meristem, SPM); 而雌穗不经历IM向BM的转变, 而是直接在IM上分化产生SPM。与BM不同, SPM具有确定性, 每个SPM产生短分枝, 并转变为小穗分生组织(spikelet meristem, SM)。SM基部产生颖壳原基, 上部形成小花分生组织(floral meristem, FM) (图2B, C)。因此, SAM向IM转变的速度以及花序发育的快慢直接决定了玉米花期长短。
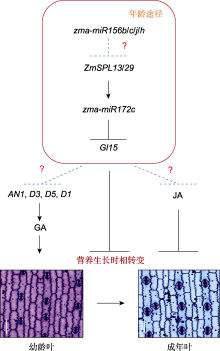
为探究调控玉米营养生长早期的遗传机制, 前人利用不同遗传背景或花期差异较大的玉米材料构建了遗传群体, 并结合分子标记图谱, 定位了一些控制营养生长时相转变的QTLs, 详细信息见表2。玉米最后一片具有蜡质的叶片是衡量玉米营养生长时相转变快慢的重要指标(Foerster et al., 2015)。基于这一表型, 进一步利用花期相近的B73与Mo17自交系构建重组自交系(recombinant inbred line, RIL)和巢式关联定位群体(nested association mapping, NAM), 通过全基因组关联分析(genome-wide association study, GWAS), 共挖掘到13个QTLs; 其中, 位于第9号染色体的SNP最为显著, 该位点对应的基因是Gl15 (Glossy15), 该基因编码AP2-like转录因子, 是zma-miR172的靶基因, 负调控营养生长时相转变(Lauter et al., 2005; Foerster et al., 2015; Xu et al., 2017); 另一个显著的SNP位于第2号染色体, 对应的基因是EDM2-like (拟南芥营养生长时相转变和成花转变关键负调控基因EDM2 (enhanced downy mildew 2)的同源基因) (Tsuchiya and Eulgem, 2010a, 2010b), 暗示EDM2-like可能参与调控玉米营养生长时相转变。此外, Xu等(2017)系统比较了长日照(long-day, LD)条件下, 玉米祖先大刍草(营养生长时相转变晚)和现代栽培玉米(营养生长时相转变早)的LLEW表型, 分别以二者典型代表8759和W22构建BC2F3群体, 再结合GWAS分析, 共鉴定到12个QTLs, 其中qVT9-1效应最大, 该位点的候选基因也对应Gl15, 其在大刍草中的表达水平远高于现代栽培玉米。进一步分析发现, Gl15的3′末端发生单核苷酸变异, 导致其在现代栽培玉米中提前终止, 表达水平降低, 从而缩短了植株的营养生长时间(Xu et al., 2017)。由此可见, 玉米营养生长时相转变由单个主效基因和多个微效基因共同调控, 其中Gl15发挥主要作用。因此, 利用营养生长时相转变差异显著的玉米种质构建群体, 结合GWAS、转录组学和代谢组学等新技术手段, 有望更快、更准确地定位到早期营养生长时相转变的关键QTLs。
本文的其它图/表
|
 首页
首页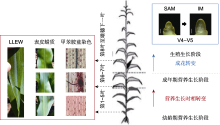
{kind=link}