水稻叶色调控机制及相关基因研究进展
戴若惠, 钱心妤, 孙静蕾, 芦涛, 贾绮玮, 陆天麒, 路梅, 饶玉春
植物学报
2023, 58 ( 5):
799-812.
DOI: 10.11983/CBB23055
植物光合作用主要依赖绿色叶片, 而叶片生长发育最直观的特征是叶色。目前已克隆200多个调控水稻(Oryza sativa)叶色的基因。水稻叶色调控机制复杂多样, 涉及多条调控途径, 包括光合色素的生物合成与降解、核-质信号转导和血红素的合成。此外, 温度和光照强度等外部环境也会影响水稻叶色的变化。该文从分子机制、叶色相关基因及环境因素等方面总结了水稻叶色遗传调控机制, 并提出该领域亟待解决的科学问题, 以期为水稻高光效育种及应用提供理论支撑。
View image in article
图3
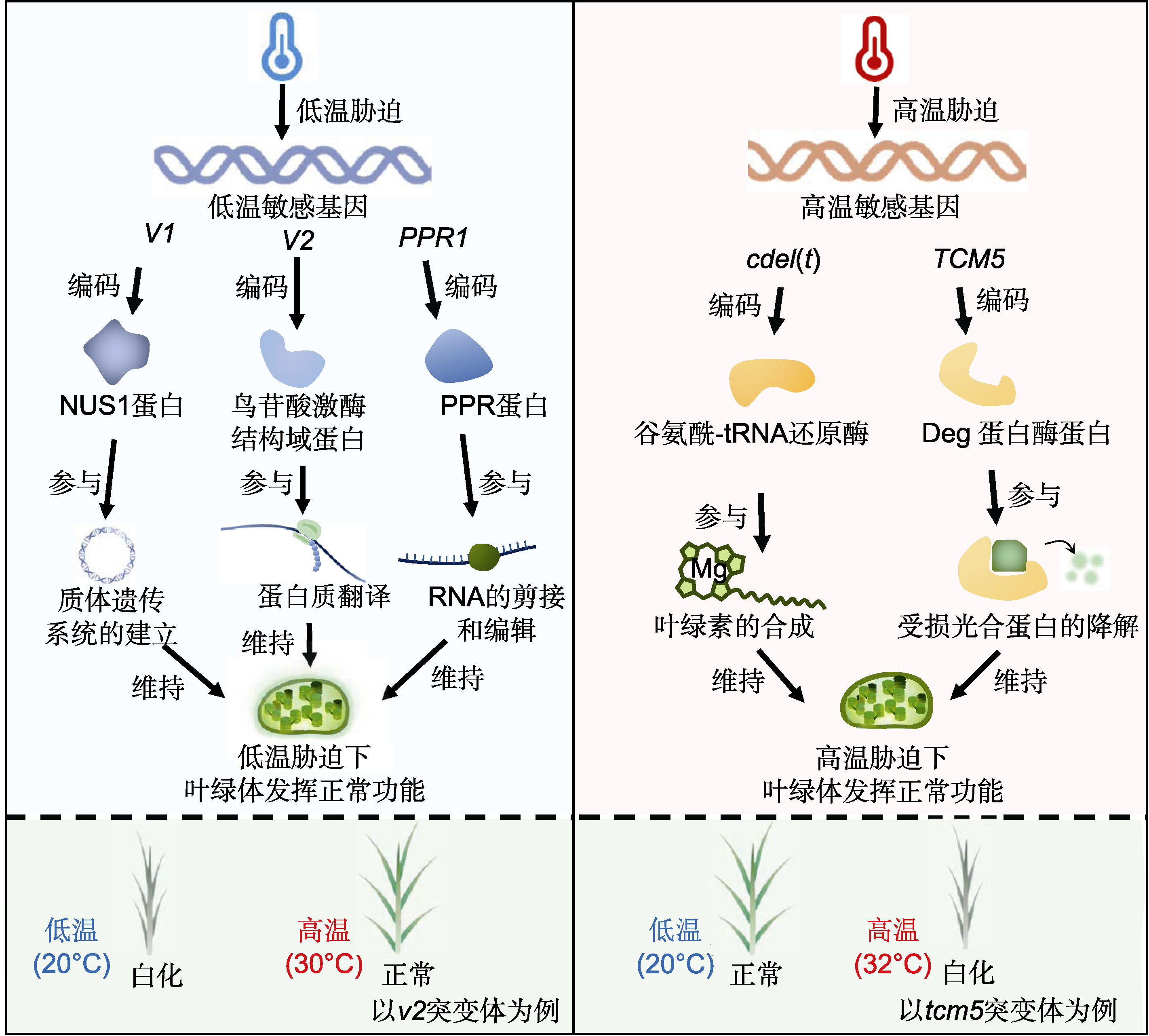
温度对水稻叶色的影响
正文中引用本图/表的段落
此外, 温度异常也会对水稻叶绿体生长发育产生影响(图3)。在低温条件下, 叶绿体内参与生理生化反应的各种酶活性降低, 使光合色素合成受抑制, 最终导致植株不能正常进行光合作用。研究发现, OsPPR1 (Gothandam et al., 2005)、OsPPR6 (Tang et al., 2017)、TCD10 (Wu et al., 2016)、DUA1 (Du et al., 2021)、OsV4 (Gong et al., 2014)和CDE4 (Liu et al., 2021b)编码PPR, PPR参与RNA的剪接和编辑(Legen et al., 2018)。这类基因突变影响基因的转录, 导致低温下水稻白化。而低温敏感基因通过调控基因转录水平和蛋白表达量响应低温胁迫。V1 (Kusumi et al., 2011)、V2 (Sugimoto et al., 2004)和TSV3 (Lin et al., 2018)分别编码NUS1蛋白、鸟苷酸激酶和苏氨酰- tRNA合成酶。其中NUS1蛋白能促进质体遗传系统的建立, 在低温下大量积累; 鸟苷酸激酶参与叶绿体内蛋白翻译; 苏氨酰-tRNA合成酶参与催化蛋白质合成。这类基因受低温诱导, 促进相关蛋白大量积累, 维持低温胁迫下叶绿体的正常生理功能。目前, 对高温敏感叶色突变体的研究相对较少。高温敏感基因有TCM5和cdel(t)等。TCM5编码叶绿体靶向的Deg蛋白酶, 32°C高温下, tcm5突变体在四到五叶期死亡, 但在低温下叶色正常, 表明TCM5在高温下影响叶绿体发育(Zheng et al., 2016)。cdel(t)编码谷氨酰-tRNA合成酶, cdel(t)突变体在超过26°C的相对高温环境下呈现黄叶表型(Liu et al., 2007)。
叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., 2011)、质体中的ROS (Laloi et al., 2006; Lee et al., 2007)及细胞器的氧化还原信号通路(Pesaresi et al., 2007)为逆行信号通路的不同来源, 对质体逆行信号分子的性质和逆行信号的积累与叶绿体形成过程的关系尚不清楚.因此, 有必要深入研究核-质信号转导途径调控叶色变化的分子机制. ... Rice NON-YELLOW COLORING 1 is involved in light-harvesting complex II and grana degradation during leaf senescence 1 2007 ... 在植物叶片中, 叶绿素合成与降解维持动态平衡.高等植物(如水稻)的叶绿素生物降解途径复杂, 任何一个环节受阻都有可能打破降解的动态平衡, 从而改变叶色.研究发现, 高等植物体内可能不仅1条叶绿素降解途径(Scheumann et al., 1998). 其中一条分解过程是在叶绿素酶(CLH)、镁离子螯合酶、脱镁叶绿酸a加氧酶(PAO)和红色叶绿素代谢产物还原酶(RCCR)的共同作用下, 将叶绿素分解为一种降解产物, 经修饰后进入液泡, 形成叶绿素代谢产物, 再转化形成单吡咯分解产物并储存于叶肉细胞中(图2).在高等植物叶绿素降解过程中, 如果相关基因发生突变, 通常会造成叶绿素分解出现异常, 使突变体植株的叶片在成熟后不褪绿变黄, 依然保持绿色.例如, NYC1编码叶绿素b还原酶, 其突变导致水稻在成熟后一直保持绿色.NOL和NYC1在体外可以发生相互作用, 且二者均定位于水稻类囊体膜, 编码短链脱氢酶/还原酶.NOL和NYC1突变后, 叶绿素b、捕光复合体II和类囊体基粒的降解均被抑制, 说明叶片衰老过程中NOL和NYC1在叶绿素b和捕光复合体II降解过程中发挥重要作用(Kusaba et al., 2007). ... Characterization of a zebra mutant of rice with increased susceptibility to light stress 1 2000 ... 水稻叶片颜色可分为白化、白翠、黄化、黄绿、淡绿、绿白、绿黄和条纹8种类型(Awan et al., 1980).叶色受温度调控, 可将水稻分为低温敏感型、温度钝感型及高温敏感型(吴殿星等, 1999).受光照影响的叶色突变体有光依赖型和非光依赖型(Kusumi et al., 2000). ... A plastid protein NUS1 is essential for build-up of the genetic system for early chloroplast development under cold stress conditions 2 2011 ... 此外, 温度异常也会对水稻叶绿体生长发育产生影响(图3).在低温条件下, 叶绿体内参与生理生化反应的各种酶活性降低, 使光合色素合成受抑制, 最终导致植株不能正常进行光合作用.研究发现, OsPPR1 (Gothandam et al., 2005)、OsPPR6 (Tang et al., 2017)、TCD10 (Wu et al., 2016)、DUA1 (Du et al., 2021)、OsV4 (Gong et al., 2014)和CDE4 (Liu et al., 2021b)编码PPR, PPR参与RNA的剪接和编辑(Legen et al., 2018).这类基因突变影响基因的转录, 导致低温下水稻白化.而低温敏感基因通过调控基因转录水平和蛋白表达量响应低温胁迫.V1 (Kusumi et al., 2011)、V2 (Sugimoto et al., 2004)和TSV3 (Lin et al., 2018)分别编码NUS1蛋白、鸟苷酸激酶和苏氨酰- tRNA合成酶.其中NUS1蛋白能促进质体遗传系统的建立, 在低温下大量积累; 鸟苷酸激酶参与叶绿体内蛋白翻译; 苏氨酰-tRNA合成酶参与催化蛋白质合成.这类基因受低温诱导, 促进相关蛋白大量积累, 维持低温胁迫下叶绿体的正常生理功能.目前, 对高温敏感叶色突变体的研究相对较少.高温敏感基因有TCM5和cdel(t)等.TCM5编码叶绿体靶向的Deg蛋白酶, 32°C高温下, tcm5突变体在四到五叶期死亡, 但在低温下叶色正常, 表明TCM5在高温下影响叶绿体发育(Zheng et al., 2016).cdel(t)编码谷氨酰-tRNA合成酶, cdel(t)突变体在超过26°C的相对高温环境下呈现黄叶表型(Liu et al., 2007). ...
叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., 2011)、质体中的ROS (Laloi et al., 2006; Lee et al., 2007)及细胞器的氧化还原信号通路(Pesaresi et al., 2007)为逆行信号通路的不同来源, 对质体逆行信号分子的性质和逆行信号的积累与叶绿体形成过程的关系尚不清楚.因此, 有必要深入研究核-质信号转导途径调控叶色变化的分子机制. ... grc1) is required for the biosynthesis of chlorophyll and the early development of chloroplasts in rice 1 2013 ... 参与血红素生物合成和降解的基因突变也会引起水稻叶色异常.血红素的生物合成与叶绿素合成的前期相同, 共用谷氨酰-tRNA到原卟啉IX途径, 之后原卟啉IX与Fe2+螯合形成血红素, 再经过一系列酶反应, 最终生成光敏色素(Tanaka et al., 2011).作为血红素到光敏色素前体转化的关键酶, 血红素加氧酶(HO)严格调控血红素的生物降解速率.Li等(2013)利用T-DNA插入得到1个关于编码血红素加氧酶基因YGL2的突变体, 突变体中代谢产物5-氨基乙酰丙酸和Mg-原卟啉IX含量升高, 而叶绿素和光敏色素前体含量降低, 导致黄叶表型. ... Mutation of the rice ASL2 gene encoding plastid ribosomal protein L21 causes chloroplast developmental defects and seedling death 1 2015a ... 质体核糖体蛋白(PRPs)是70s质体核糖体(plastori- bosomes, PR)的主要成分, 编码PRPs的基因发生突变可导致质体核糖体结构异常(Qiu et al., 2018b), 影响核-质基因转录翻译, 从而调控叶绿素含量和叶绿体发育.质体核糖体的大亚基包含33种蛋白质, 小亚基包含25种蛋白质(Yamaguchi and Subramanian, 2000).目前已对PRPs编码基因进行了大量研究, 表明PRPL3、WLP1、PRPL18、ASL2、ASL4、TCD11、WGL2和ASL1参与编码PRPs (Gong et al., 2013; Song et al., 2014; Lin et al., 2015a; Wang et al., 2017a; Qiu et al., 2018a; Lee et al., 2019; Chen et al., 2022a), 分别编码核糖体大亚基蛋白L3、L13、L18和L21及小亚基蛋白S1、S6、S9和S20. ... The rice ALS3 encoding a novel pentatricopeptide repeat protein is required for chloroplast development and seedling growth 1 2015b ... 水稻叶色表型大多由单隐性核基因控制, 如突变体als3幼苗白化致死表型(Lin et al., 2015b)、突变体vpa1白化转绿表型(王中豪等, 2021)及突变体wsl3绿化表型(Wang et al., 2016b), 但也有少数叶色是显性性状.细胞核遗传突变、细胞质遗传突变以及核-质基因互作遗传突变是水稻叶色变异的主要遗传机理.目前发现的胞质遗传较少, 仅有钱前等(1996)发现的白绿苗突变体是由细胞质基因突变导致. ... Rice TSV3 encoding obg-like GTPase protein is essential for chloroplast development during the early leaf stage under cold stress 3 2018 ... 此外, 温度异常也会对水稻叶绿体生长发育产生影响(图3).在低温条件下, 叶绿体内参与生理生化反应的各种酶活性降低, 使光合色素合成受抑制, 最终导致植株不能正常进行光合作用.研究发现, OsPPR1 (Gothandam et al., 2005)、OsPPR6 (Tang et al., 2017)、TCD10 (Wu et al., 2016)、DUA1 (Du et al., 2021)、OsV4 (Gong et al., 2014)和CDE4 (Liu et al., 2021b)编码PPR, PPR参与RNA的剪接和编辑(Legen et al., 2018).这类基因突变影响基因的转录, 导致低温下水稻白化.而低温敏感基因通过调控基因转录水平和蛋白表达量响应低温胁迫.V1 (Kusumi et al., 2011)、V2 (Sugimoto et al., 2004)和TSV3 (Lin et al., 2018)分别编码NUS1蛋白、鸟苷酸激酶和苏氨酰- tRNA合成酶.其中NUS1蛋白能促进质体遗传系统的建立, 在低温下大量积累; 鸟苷酸激酶参与叶绿体内蛋白翻译; 苏氨酰-tRNA合成酶参与催化蛋白质合成.这类基因受低温诱导, 促进相关蛋白大量积累, 维持低温胁迫下叶绿体的正常生理功能.目前, 对高温敏感叶色突变体的研究相对较少.高温敏感基因有TCM5和cdel(t)等.TCM5编码叶绿体靶向的Deg蛋白酶, 32°C高温下, tcm5突变体在四到五叶期死亡, 但在低温下叶色正常, 表明TCM5在高温下影响叶绿体发育(Zheng et al., 2016).cdel(t)编码谷氨酰-tRNA合成酶, cdel(t)突变体在超过26°C的相对高温环境下呈现黄叶表型(Liu et al., 2007). ...
叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., 2011)、质体中的ROS (Laloi et al., 2006; Lee et al., 2007)及细胞器的氧化还原信号通路(Pesaresi et al., 2007)为逆行信号通路的不同来源, 对质体逆行信号分子的性质和逆行信号的积累与叶绿体形成过程的关系尚不清楚.因此, 有必要深入研究核-质信号转导途径调控叶色变化的分子机制. ... Identification and fine mapping of a thermo-sensitive chlorophyll deficient mutant in rice (Oryza sativa L.) 1 2007 ... 此外, 温度异常也会对水稻叶绿体生长发育产生影响(图3).在低温条件下, 叶绿体内参与生理生化反应的各种酶活性降低, 使光合色素合成受抑制, 最终导致植株不能正常进行光合作用.研究发现, OsPPR1 (Gothandam et al., 2005)、OsPPR6 (Tang et al., 2017)、TCD10 (Wu et al., 2016)、DUA1 (Du et al., 2021)、OsV4 (Gong et al., 2014)和CDE4 (Liu et al., 2021b)编码PPR, PPR参与RNA的剪接和编辑(Legen et al., 2018).这类基因突变影响基因的转录, 导致低温下水稻白化.而低温敏感基因通过调控基因转录水平和蛋白表达量响应低温胁迫.V1 (Kusumi et al., 2011)、V2 (Sugimoto et al., 2004)和TSV3 (Lin et al., 2018)分别编码NUS1蛋白、鸟苷酸激酶和苏氨酰- tRNA合成酶.其中NUS1蛋白能促进质体遗传系统的建立, 在低温下大量积累; 鸟苷酸激酶参与叶绿体内蛋白翻译; 苏氨酰-tRNA合成酶参与催化蛋白质合成.这类基因受低温诱导, 促进相关蛋白大量积累, 维持低温胁迫下叶绿体的正常生理功能.目前, 对高温敏感叶色突变体的研究相对较少.高温敏感基因有TCM5和cdel(t)等.TCM5编码叶绿体靶向的Deg蛋白酶, 32°C高温下, tcm5突变体在四到五叶期死亡, 但在低温下叶色正常, 表明TCM5在高温下影响叶绿体发育(Zheng et al., 2016).cdel(t)编码谷氨酰-tRNA合成酶, cdel(t)突变体在超过26°C的相对高温环境下呈现黄叶表型(Liu et al., 2007). ... CDE4 encodes a pentatricopeptide repeat protein involved in chloroplast RNA splicing and affects chloroplast development under low-temperature conditions in rice 1 2021b ... 此外, 温度异常也会对水稻叶绿体生长发育产生影响(图3).在低温条件下, 叶绿体内参与生理生化反应的各种酶活性降低, 使光合色素合成受抑制, 最终导致植株不能正常进行光合作用.研究发现, OsPPR1 (Gothandam et al., 2005)、OsPPR6 (Tang et al., 2017)、TCD10 (Wu et al., 2016)、DUA1 (Du et al., 2021)、OsV4 (Gong et al., 2014)和CDE4 (Liu et al., 2021b)编码PPR, PPR参与RNA的剪接和编辑(Legen et al., 2018).这类基因突变影响基因的转录, 导致低温下水稻白化.而低温敏感基因通过调控基因转录水平和蛋白表达量响应低温胁迫.V1 (Kusumi et al., 2011)、V2 (Sugimoto et al., 2004)和TSV3 (Lin et al., 2018)分别编码NUS1蛋白、鸟苷酸激酶和苏氨酰- tRNA合成酶.其中NUS1蛋白能促进质体遗传系统的建立, 在低温下大量积累; 鸟苷酸激酶参与叶绿体内蛋白翻译; 苏氨酰-tRNA合成酶参与催化蛋白质合成.这类基因受低温诱导, 促进相关蛋白大量积累, 维持低温胁迫下叶绿体的正常生理功能.目前, 对高温敏感叶色突变体的研究相对较少.高温敏感基因有TCM5和cdel(t)等.TCM5编码叶绿体靶向的Deg蛋白酶, 32°C高温下, tcm5突变体在四到五叶期死亡, 但在低温下叶色正常, 表明TCM5在高温下影响叶绿体发育(Zheng et al., 2016).cdel(t)编码谷氨酰-tRNA合成酶, cdel(t)突变体在超过26°C的相对高温环境下呈现黄叶表型(Liu et al., 2007). ... OsSLC1 encodes a pentatricopeptide repeat protein essential for early chloroplast development and seedling survival 2 2020 ... Partial leaf color regulation related genes in rice3
叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., 2011)、质体中的ROS (Laloi et al., 2006; Lee et al., 2007)及细胞器的氧化还原信号通路(Pesaresi et al., 2007)为逆行信号通路的不同来源, 对质体逆行信号分子的性质和逆行信号的积累与叶绿体形成过程的关系尚不清楚.因此, 有必要深入研究核-质信号转导途径调控叶色变化的分子机制. ... WHITE PANICLE 1, a val-tRNA synthetase regulating chloroplast ribosome biogenesis in rice, is essential for early chloroplast development 1 2016b ... 水稻叶色表型大多由单隐性核基因控制, 如突变体als3幼苗白化致死表型(Lin et al., 2015b)、突变体vpa1白化转绿表型(王中豪等, 2021)及突变体wsl3绿化表型(Wang et al., 2016b), 但也有少数叶色是显性性状.细胞核遗传突变、细胞质遗传突变以及核-质基因互作遗传突变是水稻叶色变异的主要遗传机理.目前发现的胞质遗传较少, 仅有钱前等(1996)发现的白绿苗突变体是由细胞质基因突变导致. ... White panicle 2 encoding thioredoxin z, regulates plastid RNA editing by interacting with multiple organellar RNA editing factors in rice 1 2021 ... 最新研究表明, 通过对高温敏感型白化突变体wp2的研究, 证实硫氧还蛋白z通过控制细胞器RNA编辑因子(MORFs)的氧化还原状态调节植物叶绿体RNA编辑(Wang et al., 2021).研究发现五肽重复(PPR)结构域GUN1和MORF2在逆行信号转导中具有部分重叠的调控途径.GUN1也参与逆行信号转导和质体RNA编辑.gun1突变体在逆行信号转导期间调节质体中约1/3RNA编辑位点的编辑效率, 影响负责叶绿体编码的ClpP的转录效率.其中, ClpP的RNA编辑水平变化可能最终影响Clp蛋白酶活性, 导致未折叠蛋白质聚集(Zhao et al., 2019).该发现在叶绿体逆行信号与核RNA代谢过程或质体RNA编辑联系方面取得了实质性进展.OsPPR6也参与编辑和剪接叶绿体RNA的PPR, 为水稻叶绿体生物发生所必需.Tang等(2017)研究发现osppr6突变体中质体编码的RNA聚合酶减少, 而核编码的RNA聚合酶转录的基因增多, 经验证OsPPR6特异性编辑叶绿体ndhB转录物, 是转录物YCF3 (光系统I组装蛋白)的必要物. ... Heme synthesis by plastid ferrochelatase I regulates nuclear gene expression in plants 1 2011 ... 叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., 2011)、质体中的ROS (Laloi et al., 2006; Lee et al., 2007)及细胞器的氧化还原信号通路(Pesaresi et al., 2007)为逆行信号通路的不同来源, 对质体逆行信号分子的性质和逆行信号的积累与叶绿体形成过程的关系尚不清楚.因此, 有必要深入研究核-质信号转导途径调控叶色变化的分子机制. ... The rice pentatricopeptide repeat gene TCD10 is needed for chloroplast development under cold stress 1 2016 ... 此外, 温度异常也会对水稻叶绿体生长发育产生影响(图3).在低温条件下, 叶绿体内参与生理生化反应的各种酶活性降低, 使光合色素合成受抑制, 最终导致植株不能正常进行光合作用.研究发现, OsPPR1 (Gothandam et al., 2005)、OsPPR6 (Tang et al., 2017)、TCD10 (Wu et al., 2016)、DUA1 (Du et al., 2021)、OsV4 (Gong et al., 2014)和CDE4 (Liu et al., 2021b)编码PPR, PPR参与RNA的剪接和编辑(Legen et al., 2018).这类基因突变影响基因的转录, 导致低温下水稻白化.而低温敏感基因通过调控基因转录水平和蛋白表达量响应低温胁迫.V1 (Kusumi et al., 2011)、V2 (Sugimoto et al., 2004)和TSV3 (Lin et al., 2018)分别编码NUS1蛋白、鸟苷酸激酶和苏氨酰- tRNA合成酶.其中NUS1蛋白能促进质体遗传系统的建立, 在低温下大量积累; 鸟苷酸激酶参与叶绿体内蛋白翻译; 苏氨酰-tRNA合成酶参与催化蛋白质合成.这类基因受低温诱导, 促进相关蛋白大量积累, 维持低温胁迫下叶绿体的正常生理功能.目前, 对高温敏感叶色突变体的研究相对较少.高温敏感基因有TCM5和cdel(t)等.TCM5编码叶绿体靶向的Deg蛋白酶, 32°C高温下, tcm5突变体在四到五叶期死亡, 但在低温下叶色正常, 表明TCM5在高温下影响叶绿体发育(Zheng et al., 2016).cdel(t)编码谷氨酰-tRNA合成酶, cdel(t)突变体在超过26°C的相对高温环境下呈现黄叶表型(Liu et al., 2007). ... A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis 2 2007 ... 叶绿素合成是从L-谷氨酰-tRNA开始到叶绿素a, 再到叶绿素b.该过程主要分4个步骤.第1步, 谷氨酰- tRNA经谷氨酰-tRNA合成酶、谷氨酰-tRNA还原酶和谷氨酸-1-半醛氨基酸转移酶的催化以及一系列反应形成5-氨基酮戊酸(ALA), ALA是血红素和叶绿素的共同前体, 当植物中血红素代谢受阻导致血红素含量增加时, 通过负反馈调节抑制ALA合成, 从而抑制叶绿素的合成, 最终导致叶色改变(Terry and Kendrick, 1999).第2步, ALA经过5-氨基酮戊酸脱水酶、胆色素原脱氨酶以及尿卟啉原III合成酶等一系列催化反应形成尿卟啉原III.第3步, 尿卟啉原III在叶绿体基质中经过粪卟啉原III氧化酶和原卟啉原氧化酶的催化形成原卟啉IX.第4步, 原卟啉IX与镁离子螯合, 经过多个膜结合酶催化反应生成叶绿素a, 在叶绿素a加氧酶和柠檬酸合酶(CS)的催化下形成叶绿素b (Matringe et al., 1992; Von Wettstein et al., 1995; Wu et al., 2007) (图1).整个过程有20多个基因编码的15种酶参与, 其中任何一个生物化学步骤涉及的基因或酶变异都有可能导致叶色异常.在合成途径中, 受阻发生得越早, 叶色突变越明显, 一般表现为白化或黄化; 如果发生在合成后期, 通常只会表现为条纹或斑块(Sakuraba et al., 2012). ...
叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., 2011)、质体中的ROS (Laloi et al., 2006; Lee et al., 2007)及细胞器的氧化还原信号通路(Pesaresi et al., 2007)为逆行信号通路的不同来源, 对质体逆行信号分子的性质和逆行信号的积累与叶绿体形成过程的关系尚不清楚.因此, 有必要深入研究核-质信号转导途径调控叶色变化的分子机制. ... GUN1 interacts with MORF2 to regulate plastid RNA editing during retrograde signaling 1 2019 ... 最新研究表明, 通过对高温敏感型白化突变体wp2的研究, 证实硫氧还蛋白z通过控制细胞器RNA编辑因子(MORFs)的氧化还原状态调节植物叶绿体RNA编辑(Wang et al., 2021).研究发现五肽重复(PPR)结构域GUN1和MORF2在逆行信号转导中具有部分重叠的调控途径.GUN1也参与逆行信号转导和质体RNA编辑.gun1突变体在逆行信号转导期间调节质体中约1/3RNA编辑位点的编辑效率, 影响负责叶绿体编码的ClpP的转录效率.其中, ClpP的RNA编辑水平变化可能最终影响Clp蛋白酶活性, 导致未折叠蛋白质聚集(Zhao et al., 2019).该发现在叶绿体逆行信号与核RNA代谢过程或质体RNA编辑联系方面取得了实质性进展.OsPPR6也参与编辑和剪接叶绿体RNA的PPR, 为水稻叶绿体生物发生所必需.Tang等(2017)研究发现osppr6突变体中质体编码的RNA聚合酶减少, 而核编码的RNA聚合酶转录的基因增多, 经验证OsPPR6特异性编辑叶绿体ndhB转录物, 是转录物YCF3 (光系统I组装蛋白)的必要物. ... The rice TCM5 gene encoding a novel deg protease protein is essential for chloroplast development under high temperatures 1 2016 ... 此外, 温度异常也会对水稻叶绿体生长发育产生影响(图3).在低温条件下, 叶绿体内参与生理生化反应的各种酶活性降低, 使光合色素合成受抑制, 最终导致植株不能正常进行光合作用.研究发现, OsPPR1 (Gothandam et al., 2005)、OsPPR6 (Tang et al., 2017)、TCD10 (Wu et al., 2016)、DUA1 (Du et al., 2021)、OsV4 (Gong et al., 2014)和CDE4 (Liu et al., 2021b)编码PPR, PPR参与RNA的剪接和编辑(Legen et al., 2018).这类基因突变影响基因的转录, 导致低温下水稻白化.而低温敏感基因通过调控基因转录水平和蛋白表达量响应低温胁迫.V1 (Kusumi et al., 2011)、V2 (Sugimoto et al., 2004)和TSV3 (Lin et al., 2018)分别编码NUS1蛋白、鸟苷酸激酶和苏氨酰- tRNA合成酶.其中NUS1蛋白能促进质体遗传系统的建立, 在低温下大量积累; 鸟苷酸激酶参与叶绿体内蛋白翻译; 苏氨酰-tRNA合成酶参与催化蛋白质合成.这类基因受低温诱导, 促进相关蛋白大量积累, 维持低温胁迫下叶绿体的正常生理功能.目前, 对高温敏感叶色突变体的研究相对较少.高温敏感基因有TCM5和cdel(t)等.TCM5编码叶绿体靶向的Deg蛋白酶, 32°C高温下, tcm5突变体在四到五叶期死亡, 但在低温下叶色正常, 表明TCM5在高温下影响叶绿体发育(Zheng et al., 2016).cdel(t)编码谷氨酰-tRNA合成酶, cdel(t)突变体在超过26°C的相对高温环境下呈现黄叶表型(Liu et al., 2007). ... C-terminal residues of Oryza sativa GUN4 are required for the activation of the ChlH subunit of magnesium chelatase in chlorophyll synthesis 1 2012 ... Partial leaf color regulation related genes in rice3
本文的其它图/表
|

 首页
首页
{kind=link}