
|
|
||
|
水稻叶色调控机制及相关基因研究进展
植物学报
2023, 58 (5):
799-812.
DOI: 10.11983/CBB23055
植物光合作用主要依赖绿色叶片, 而叶片生长发育最直观的特征是叶色。目前已克隆200多个调控水稻(Oryza sativa)叶色的基因。水稻叶色调控机制复杂多样, 涉及多条调控途径, 包括光合色素的生物合成与降解、核-质信号转导和血红素的合成。此外, 温度和光照强度等外部环境也会影响水稻叶色的变化。该文从分子机制、叶色相关基因及环境因素等方面总结了水稻叶色遗传调控机制, 并提出该领域亟待解决的科学问题, 以期为水稻高光效育种及应用提供理论支撑。
 View image in article
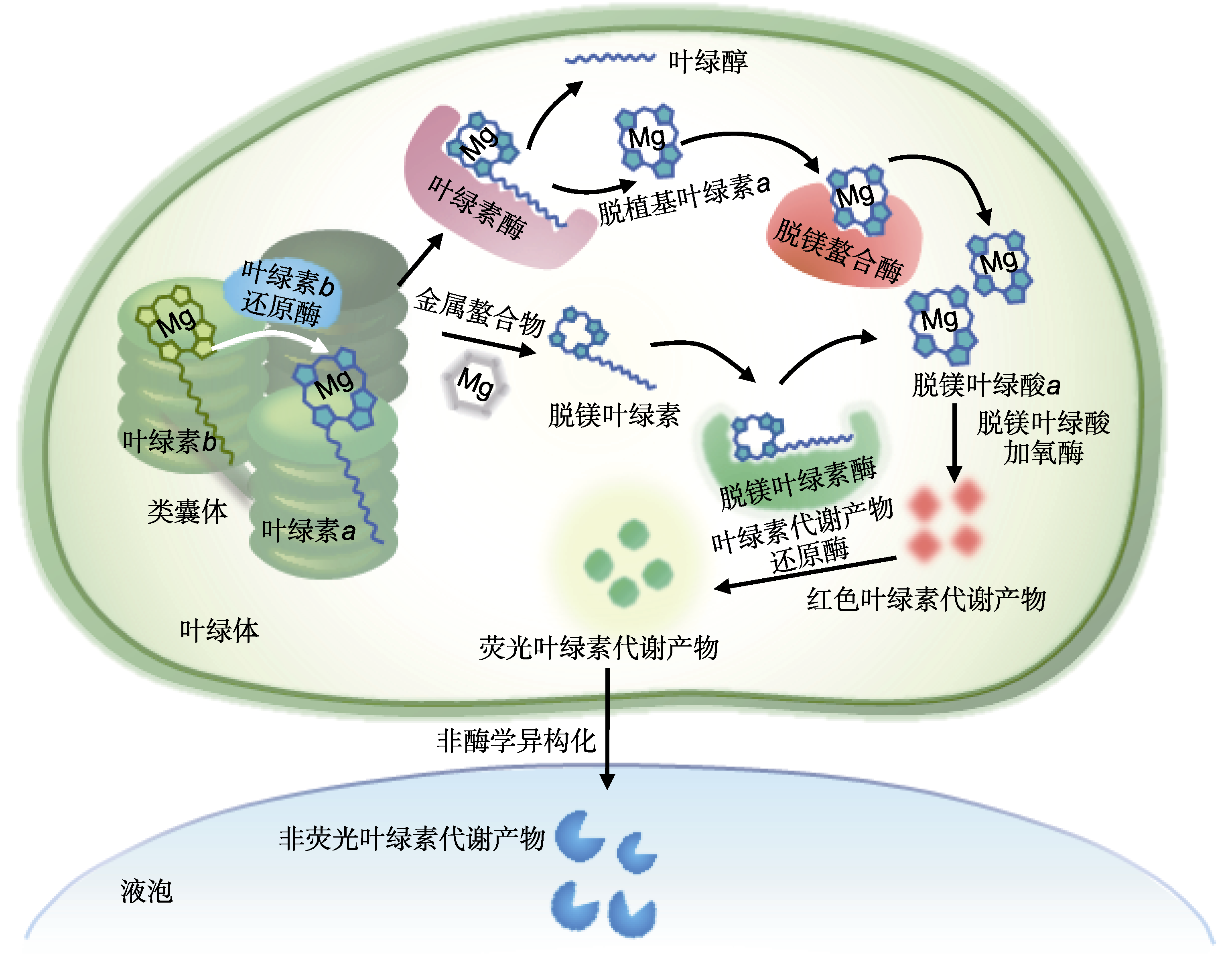
图2
叶绿素在叶绿体内的降解途径
正文中引用本图/表的段落
在植物叶片中, 叶绿素合成与降解维持动态平衡。高等植物(如水稻)的叶绿素生物降解途径复杂, 任何一个环节受阻都有可能打破降解的动态平衡, 从而改变叶色。研究发现, 高等植物体内可能不仅1条叶绿素降解途径(Scheumann et al., 1998)。 其中一条分解过程是在叶绿素酶(CLH)、镁离子螯合酶、脱镁叶绿酸a加氧酶(PAO)和红色叶绿素代谢产物还原酶(RCCR)的共同作用下, 将叶绿素分解为一种降解产物, 经修饰后进入液泡, 形成叶绿素代谢产物, 再转化形成单吡咯分解产物并储存于叶肉细胞中(图2)。在高等植物叶绿素降解过程中, 如果相关基因发生突变, 通常会造成叶绿素分解出现异常, 使突变体植株的叶片在成熟后不褪绿变黄, 依然保持绿色。例如, NYC1编码叶绿素b还原酶, 其突变导致水稻在成熟后一直保持绿色。NOL和NYC1在体外可以发生相互作用, 且二者均定位于水稻类囊体膜, 编码短链脱氢酶/还原酶。NOL和NYC1突变后, 叶绿素b、捕光复合体II和类囊体基粒的降解均被抑制, 说明叶片衰老过程中NOL和NYC1在叶绿素b和捕光复合体II降解过程中发挥重要作用(Kusaba et al., 2007)。
叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., Rice NON-YELLOW COLORING 1 is involved in light-harvesting complex II and grana degradation during leaf senescence 1 2007 ... 在植物叶片中, 叶绿素合成与降解维持动态平衡.高等植物(如水稻)的叶绿素生物降解途径复杂, 任何一个环节受阻都有可能打破降解的动态平衡, 从而改变叶色.研究发现, 高等植物体内可能不仅1条叶绿素降解途径(Scheumann et al., Characterization of a zebra mutant of rice with increased susceptibility to light stress 1 2000 ... 水稻叶片颜色可分为白化、白翠、黄化、黄绿、淡绿、绿白、绿黄和条纹8种类型(Awan et al., A plastid protein NUS1 is essential for build-up of the genetic system for early chloroplast development under cold stress conditions 2 2011 ... 此外, 温度异常也会对水稻叶绿体生长发育产生影响(
叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., STAY-GREEN and chlorophyll catabolic enzymes interact at light-harvesting complex II for chlorophyll detoxification during leaf senescence in Arabidopsis 1 2012 ... 叶绿素合成是从L-谷氨酰-tRNA开始到叶绿素a, 再到叶绿素b.该过程主要分4个步骤.第1步, 谷氨酰- tRNA经谷氨酰-tRNA合成酶、谷氨酰-tRNA还原酶和谷氨酸-1-半醛氨基酸转移酶的催化以及一系列反应形成5-氨基酮戊酸(ALA), ALA是血红素和叶绿素的共同前体, 当植物中血红素代谢受阻导致血红素含量增加时, 通过负反馈调节抑制ALA合成, 从而抑制叶绿素的合成, 最终导致叶色改变(Terry and Kendrick, Chlorophyll a formation in the chlorophyll b reductase reaction requires reduced ferredoxin 1 1998 ... 在植物叶片中, 叶绿素合成与降解维持动态平衡.高等植物(如水稻)的叶绿素生物降解途径复杂, 任何一个环节受阻都有可能打破降解的动态平衡, 从而改变叶色.研究发现, 高等植物体内可能不仅1条叶绿素降解途径(Scheumann et al., The rice nuclear gene WLP1 encoding a chloroplast ribosome L13 protein is needed for chloroplast development in rice grown under low temperature conditions 3 2014 ... 质体核糖体蛋白(PRPs)是70s质体核糖体(plastori- bosomes, PR)的主要成分, 编码PRPs的基因发生突变可导致质体核糖体结构异常(Qiu et al.,
本文的其它图/表
|
 首页
首页
{kind=link}