
|
|
||
|
水稻叶色调控机制及相关基因研究进展
植物学报
2023, 58 (5):
799-812.
DOI: 10.11983/CBB23055
植物光合作用主要依赖绿色叶片, 而叶片生长发育最直观的特征是叶色。目前已克隆200多个调控水稻(Oryza sativa)叶色的基因。水稻叶色调控机制复杂多样, 涉及多条调控途径, 包括光合色素的生物合成与降解、核-质信号转导和血红素的合成。此外, 温度和光照强度等外部环境也会影响水稻叶色的变化。该文从分子机制、叶色相关基因及环境因素等方面总结了水稻叶色遗传调控机制, 并提出该领域亟待解决的科学问题, 以期为水稻高光效育种及应用提供理论支撑。
 View image in article
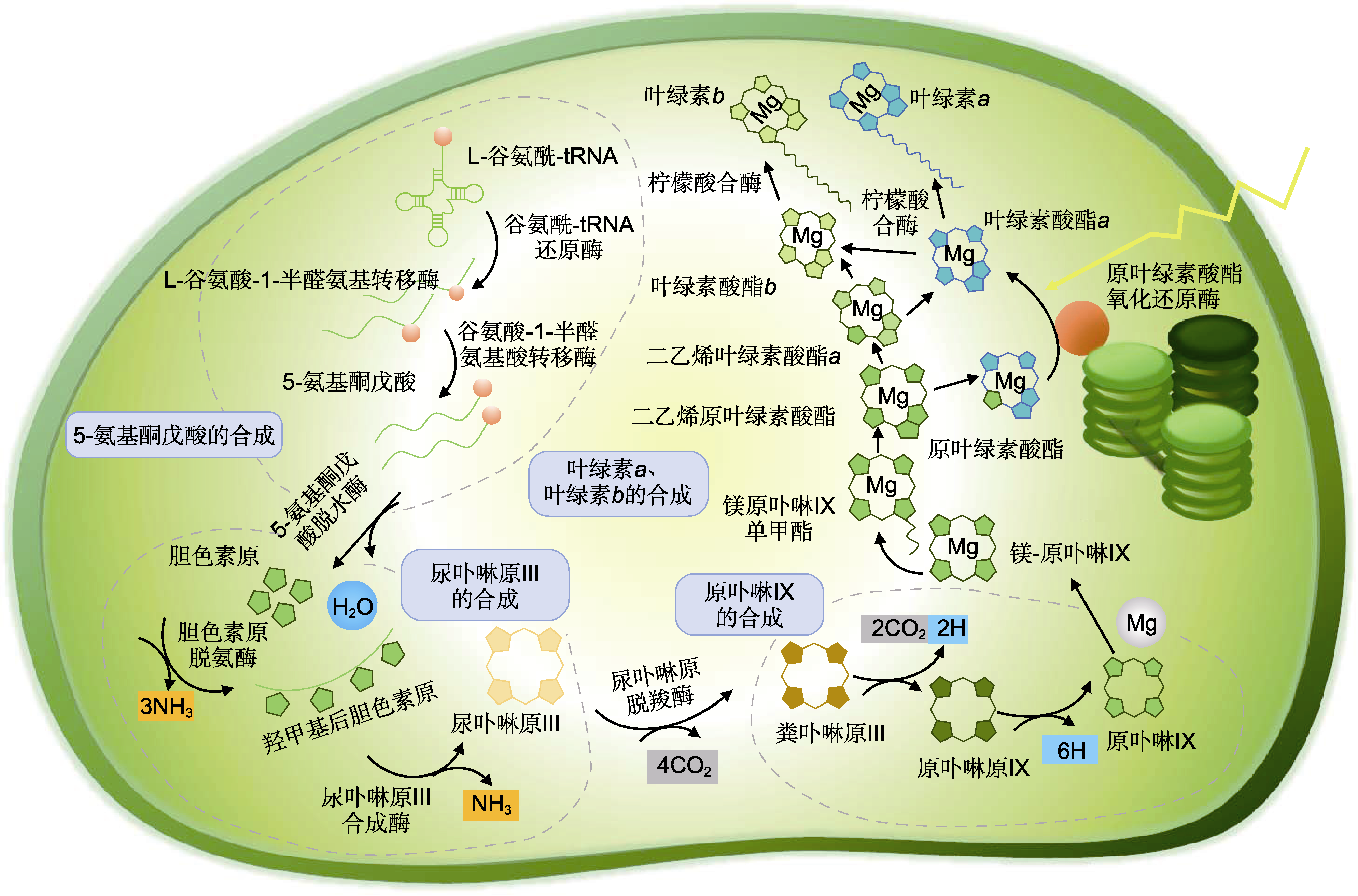
图1
叶绿素在叶绿体内的合成途径
正文中引用本图/表的段落
叶绿素合成是从L-谷氨酰-tRNA开始到叶绿素a, 再到叶绿素b。该过程主要分4个步骤。第1步, 谷氨酰- tRNA经谷氨酰-tRNA合成酶、谷氨酰-tRNA还原酶和谷氨酸-1-半醛氨基酸转移酶的催化以及一系列反应形成5-氨基酮戊酸(ALA), ALA是血红素和叶绿素的共同前体, 当植物中血红素代谢受阻导致血红素含量增加时, 通过负反馈调节抑制ALA合成, 从而抑制叶绿素的合成, 最终导致叶色改变(Terry and Kendrick, 1999)。第2步, ALA经过5-氨基酮戊酸脱水酶、胆色素原脱氨酶以及尿卟啉原III合成酶等一系列催化反应形成尿卟啉原III。第3步, 尿卟啉原III在叶绿体基质中经过粪卟啉原III氧化酶和原卟啉原氧化酶的催化形成原卟啉IX。第4步, 原卟啉IX与镁离子螯合, 经过多个膜结合酶催化反应生成叶绿素a, 在叶绿素a加氧酶和柠檬酸合酶(CS)的催化下形成叶绿素b (Matringe et al., 1992; Von Wettstein et al., 1995; Wu et al., 2007) (图1)。整个过程有20多个基因编码的15种酶参与, 其中任何一个生物化学步骤涉及的基因或酶变异都有可能导致叶色异常。在合成途径中, 受阻发生得越早, 叶色突变越明显, 一般表现为白化或黄化; 如果发生在合成后期, 通常只会表现为条纹或斑块(Sakuraba et al., 2012)。
叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., Localization within chloroplasts of protoporphyrinogen oxidase, the target enzyme for diphenylether-like herbicides 1 1992 ... 叶绿素合成是从L-谷氨酰-tRNA开始到叶绿素a, 再到叶绿素b.该过程主要分4个步骤.第1步, 谷氨酰- tRNA经谷氨酰-tRNA合成酶、谷氨酰-tRNA还原酶和谷氨酸-1-半醛氨基酸转移酶的催化以及一系列反应形成5-氨基酮戊酸(ALA), ALA是血红素和叶绿素的共同前体, 当植物中血红素代谢受阻导致血红素含量增加时, 通过负反馈调节抑制ALA合成, 从而抑制叶绿素的合成, 最终导致叶色改变(Terry and Kendrick, Newly identified CSP41b gene localized in chloroplasts affects leaf color in rice 1 2017 ... 叶绿体内的类囊体膜是植物色素代谢的主要场所, 其内部结构的变化与叶色异常密切相关.研究表明, 相比正常绿叶植物, 叶色变异植物的叶绿体形态和结构有很大差异, 具体表现为叶绿体外膜不清晰、不完整, 叶绿体中类囊体基质质地稀薄, 基粒类囊体数量偏少以及片层垛叠稀疏, 基因突变程度较高者甚至很难分化出完整的类囊体结构(孙旺旺, Identification of the carotenoid isomerase provides insight into carotenoid biosynthesis, prolamellar body formation, and photomorphogenesis 1 2002 ... 目前已知类胡萝卜素含量变化主要通过调控合成途径中酶基因的表达实现(樊宝莲和王晓云, Interorganellar communication 1 2007 ... 叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., The rice white green leaf 2 gene causes defects in chloroplast development and affects the plastid ribosomal protein S9 2 2018a ... 质体核糖体蛋白(PRPs)是70s质体核糖体(plastori- bosomes, PR)的主要成分, 编码PRPs的基因发生突变可导致质体核糖体结构异常(Qiu et al.,
叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., STAY-GREEN and chlorophyll catabolic enzymes interact at light-harvesting complex II for chlorophyll detoxification during leaf senescence in Arabidopsis 1 2012 ... 叶绿素合成是从L-谷氨酰-tRNA开始到叶绿素a, 再到叶绿素b.该过程主要分4个步骤.第1步, 谷氨酰- tRNA经谷氨酰-tRNA合成酶、谷氨酰-tRNA还原酶和谷氨酸-1-半醛氨基酸转移酶的催化以及一系列反应形成5-氨基酮戊酸(ALA), ALA是血红素和叶绿素的共同前体, 当植物中血红素代谢受阻导致血红素含量增加时, 通过负反馈调节抑制ALA合成, 从而抑制叶绿素的合成, 最终导致叶色改变(Terry and Kendrick, Chlorophyll a formation in the chlorophyll b reductase reaction requires reduced ferredoxin 1 1998 ... 在植物叶片中, 叶绿素合成与降解维持动态平衡.高等植物(如水稻)的叶绿素生物降解途径复杂, 任何一个环节受阻都有可能打破降解的动态平衡, 从而改变叶色.研究发现, 高等植物体内可能不仅1条叶绿素降解途径(Scheumann et al., The rice nuclear gene WLP1 encoding a chloroplast ribosome L13 protein is needed for chloroplast development in rice grown under low temperature conditions 3 2014 ... 质体核糖体蛋白(PRPs)是70s质体核糖体(plastori- bosomes, PR)的主要成分, 编码PRPs的基因发生突变可导致质体核糖体结构异常(Qiu et al.,
叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., Feedback inhibition of chlorophyll synthesis in the phytochrome chromophore- deficient aurea and yellow-green-2 mutants of tomato 1 1999 ... 叶绿素合成是从L-谷氨酰-tRNA开始到叶绿素a, 再到叶绿素b.该过程主要分4个步骤.第1步, 谷氨酰- tRNA经谷氨酰-tRNA合成酶、谷氨酰-tRNA还原酶和谷氨酸-1-半醛氨基酸转移酶的催化以及一系列反应形成5-氨基酮戊酸(ALA), ALA是血红素和叶绿素的共同前体, 当植物中血红素代谢受阻导致血红素含量增加时, 通过负反馈调节抑制ALA合成, 从而抑制叶绿素的合成, 最终导致叶色改变(Terry and Kendrick, Chlorophyll biosynthesis 1 1995 ... 叶绿素合成是从L-谷氨酰-tRNA开始到叶绿素a, 再到叶绿素b.该过程主要分4个步骤.第1步, 谷氨酰- tRNA经谷氨酰-tRNA合成酶、谷氨酰-tRNA还原酶和谷氨酸-1-半醛氨基酸转移酶的催化以及一系列反应形成5-氨基酮戊酸(ALA), ALA是血红素和叶绿素的共同前体, 当植物中血红素代谢受阻导致血红素含量增加时, 通过负反馈调节抑制ALA合成, 从而抑制叶绿素的合成, 最终导致叶色改变(Terry and Kendrick, Divinyl chlorophyll(ide) a can be converted to monovinyl chlorophyll(ide) a by a divinyl reductase in rice 1 2010 ... 核基因控制叶绿体形成过程中的蛋白质加工和运输、光合作用元件的形成、叶绿素的积累和光感受器信号转导等.目前对水稻核基因OsDVR (LOC_Os03g22780)已有深入研究.OsDVR编码的二乙烯基还原酶(DVR)将各种叶绿素(Chl)中间体上的8-乙烯基基团转化为乙基基团, 包括将3,8-二乙烯基Chl (DV-Chla)转化为3-乙烯基Chl (MV-Chla)和联乙烯叶绿素酸酯a (DV-Chlide a)转化为单乙烯叶绿素酸酯a (MV-Chlide a)等5条转化途径, 这些途径为叶绿素a生物合成所必需.OsDVR失活的824ys突变体在不同发育阶段的各器官中专门积累二乙烯基叶绿素, 使叶绿体发育受阻, 导致叶色变为黄绿色(Wang et al., One divinyl reductase reduces the 8-vinyl groups in various intermediates of chlorophyll biosynthesis in a given higher plant species, but the isozyme differs between species 2 2013 ... 核基因控制叶绿体形成过程中的蛋白质加工和运输、光合作用元件的形成、叶绿素的积累和光感受器信号转导等.目前对水稻核基因OsDVR (LOC_Os03g22780)已有深入研究.OsDVR编码的二乙烯基还原酶(DVR)将各种叶绿素(Chl)中间体上的8-乙烯基基团转化为乙基基团, 包括将3,8-二乙烯基Chl (DV-Chla)转化为3-乙烯基Chl (MV-Chla)和联乙烯叶绿素酸酯a (DV-Chlide a)转化为单乙烯叶绿素酸酯a (MV-Chlide a)等5条转化途径, 这些途径为叶绿素a生物合成所必需.OsDVR失活的824ys突变体在不同发育阶段的各器官中专门积累二乙烯基叶绿素, 使叶绿体发育受阻, 导致叶色变为黄绿色(Wang et al.,
叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., WHITE PANICLE 1, a val-tRNA synthetase regulating chloroplast ribosome biogenesis in rice, is essential for early chloroplast development 1 2016b ... 水稻叶色表型大多由单隐性核基因控制, 如突变体als3幼苗白化致死表型(Lin et al., White panicle 2 encoding thioredoxin z, regulates plastid RNA editing by interacting with multiple organellar RNA editing factors in rice 1 2021 ... 最新研究表明, 通过对高温敏感型白化突变体wp2的研究, 证实硫氧还蛋白z通过控制细胞器RNA编辑因子(MORFs)的氧化还原状态调节植物叶绿体RNA编辑(Wang et al., Heme synthesis by plastid ferrochelatase I regulates nuclear gene expression in plants 1 2011 ... 叶色突变研究基础较为深厚, 但相关研究主要集中于叶绿体结构、色素合成与降解的分子机制.叶色相关的核-质转运分子机制目前仅被初步阐明, 其中质体-核逆向信号通路的研究进展缓慢, 仅明确四吡咯生物合成途径的中间体组分(Woodson et al., The rice pentatricopeptide repeat gene TCD10 is needed for chloroplast development under cold stress 1 2016 ... 此外, 温度异常也会对水稻叶绿体生长发育产生影响( A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis 2 2007 ... 叶绿素合成是从L-谷氨酰-tRNA开始到叶绿素a, 再到叶绿素b.该过程主要分4个步骤.第1步, 谷氨酰- tRNA经谷氨酰-tRNA合成酶、谷氨酰-tRNA还原酶和谷氨酸-1-半醛氨基酸转移酶的催化以及一系列反应形成5-氨基酮戊酸(ALA), ALA是血红素和叶绿素的共同前体, 当植物中血红素代谢受阻导致血红素含量增加时, 通过负反馈调节抑制ALA合成, 从而抑制叶绿素的合成, 最终导致叶色改变(Terry and Kendrick,
本文的其它图/表
|
 首页
首页
{kind=link}