植物GH3基因家族生物学功能研究进展
园园, 恩和巴雅尔, 齐艳华
植物学报
2023, 58 ( 5):
770-782.
DOI: 10.11983/CBB22263
植物生长素早期响应基因GH3编码的酰胺合酶催化生长素、茉莉酸及苯甲酸衍生物与氨基酸结合, 形成相应的氨基酸复合物。当植物体内生长素浓度过高时, GH3蛋白催化生长素与氨基酸结合, 形成的复合物作为生长素贮存库。当生长素浓度过低时, 生长素-氨基酸复合物被蛋白水解酶水解为生长素, 重新参与生长素信号通路, 从而调控植物体内生长素动态平衡。当植物受到生物或非生物胁迫时, GH3蛋白催化茉莉酸和水杨酸与氨基酸结合, 参与植物胁迫响应。该文从GH3蛋白结构、GH3基因家族分类及其生物学功能方面总结了双子叶模式植物拟南芥(Arabidopsis thaliana)、单子叶模式植物水稻(Oryza sativa)及其它植物中GH3基因的研究进展, 为植物GH3基因家族的深入研究提供参考。
| 基因编号 | 基因名称 | 功能 | 参考文献 | | At2g14960.1 | AtGH3.1 | 调节吲哚-3-乙酸(IAA)动态平衡 | Staswick et al., 2005 | | At4g37390.1 | AtGH3.2/YDK1 | 调控株高、主根伸长、侧根数与顶端优势 | Takase et al., 2004; Staswick et al., 2005 | | At4g27260.1 | AtGH3.5/AtGH3a/WES1 | 调控主根伸长及侧根数, 参与水杨酸(SA)与光信号途径 | Staswick et al., 2005; Park et al., 2007b; Zhang et al., 2007, 2008 | | At5g54510.1 | AtGH3.6/DFL1 | 调控生长素含量, 影响株高、叶形态、侧根数及非生物胁迫响应, 参与光信号途径, 影响下胚轴伸长 | Nakazawa et al., 2001; 刘晓东等, 2016 | | At2g47750.1 | AtGH3.9 | 调控株高、花器官以及果实发育 | 周苹等, 2015 | | At4g03400.1 | AtGH3.10/DFL2 | 参与光信号途径, 影响下胚轴伸长 | Takase et al., 2003 | | At2g46370.1 | AtGH3.11/JAR1/FIN219 | 调控茉莉酸(JA)水平, 影响抗病性, 参与光信号转导途径 | Staswick et al., 1998; van Loon et al., 1998; Overmyer et al., 2000; Rao et al., 2000; Hsieh et al., 2000 | | At5g13320.1 | AtGH3.12/PBS3/GDG1 | 调控SA水平, 影响植物抗病性 | Jagadeeswaran et al., 2007; Nobuta et al., 2007 | | At5g13370.1 | AtGH3.15 | 调节吲哚丁酸(IBA)水平, 影响主根伸长与侧根数 | Sherp et al., 2018 | | At1g28130.1 | AtGH3.17 | 调控株高、根系发育及叶片形态, 参与油菜素内酯(BL)信号途径 | 周淑瑶等, 2023 |
View table in article
表1
拟南芥GH3基因的生物学功能
正文中引用本图/表的段落
GH3亚家族I催化JA与氨基酸结合, 形成具有生物活性的茉莉酸异亮氨酸复合物(JA-Ile) (Staswick et al., 2002)。茉莉酸是植物体内重要的脂质激素, 在植物胁迫响应中发挥重要作用。同时, 茉莉酸作为重要的生长发育调节物质, 参与调节植物生长发育的诸多过程(Wasternack and Hause, 2013; 李梦莎和阎秀峰, 2014; 黎家和李传友, 2019)。当植物受到机械损伤或昆虫啃食时, 受伤部位迅速积累的JA被JAR1/AtGH3.11催化形成JA信号转导中的主要活性复合物JA-Ile。JA-Ile先被SCFCOI1受体识别, 后招募茉莉酸响应抑制因子JAZ (jasmonate ZIM-domain), 形成SCFCOI1-JA-Ile-JAZ三元复合物(Yan et al., 2018)。形成的三元复合物使JAZ被26S蛋白降解, 从而解除对转录因子(transcription factors, TF) MYC2的抑制作用, 激活JA响应基因的表达(图1) (李梦莎和阎秀峰, 2014)。
拟南芥20个GH3基因可分为3个亚家族。亚家族I包含AtGH3.10和AtGH3.11; 亚家族II包含AtGH3.1、AtGH3.2、AtGH3.3、AtGH3.4、AtGH3.5、AtGH3.6、AtGH3.9和AtGH3.17; 其余拟南芥GH3基因归为亚家族III (Staswick et al., 2002)。研究表明GH3基因在拟南芥生长发育及环境响应中发挥重要作用(表1; 图3)。
研究表明GH3家族基因调控拟南芥生长发育及形态建成。由AtGH3s基因表达引起的游离生长素含量变化影响拟南芥植株构型及根系发育。过表达AtGH3.6获得拟南芥dfl1-D矮化株系, 较野生型茎更短、叶更小和侧根数更少(Nakazawa et al., 2001)。YDK1/AtGH3.2过表达株系ydk1-D矮化突变体表现主根短、侧根数减少和顶端优势减弱, 并且YDK1表达受生长素响应因子ARF7调节(Takase et al., 2004)。AtGH3.5过表达株系同样表现出生长素缺陷表型, 具有更小的卷曲莲座叶, 更耐外源生长素类物质且主根缩短、侧根数减少(Zhang et al., 2007)。AtGH3.9过表达株系除了植株矮化外, 还表现出雄蕊变短和果荚短小(周苹等, 2015)。AtGH3.17过表达株系表现出短根、叶片卷曲、叶柄缩短和矮化等表型, 并且IAA-Glu含量显著增加, 油菜素内酯(BL)合成途径相关基因DWF4和CPD的表达量明显增高, 表明AtGH3.17除调节IAA含量外还参与BR信号途径(周淑瑶等, 2023)。值得注意的是, AtGH3.15过表达株系在主根伸长和侧根密度上对吲哚丁酸(IBA)处理有抗性, 但对IAA或JA处理无抗性, 说明AtGH3.15可能通过调节过氧化物酶体转化为IAA的IBA水平, 在生长素平衡中发挥作用(Sherp et al., 2018)。atgh3.1-1、atgh3.2-1、atgh3.5-1、atgh3.9和atgh3.17-1等单基因突变体可能由于存在基因功能冗余而未表现出明显的表型, 但因为突变体中生长素动态平衡调节机制受损导致主根生长对外源生长素类物质敏感(Staswick et al., 2005; Khan and Stone, 2007)。
植物在生长发育过程中常受到生物与非生物胁迫, JA和SA作为信号分子在植物胁迫响应中发挥重要作用。目前, GH3基因参与拟南芥胁迫响应的研究主要集中在少数几个AtGH3s基因中。Staswick等(1998)研究发现, JAR1/AtGH3.11突变体jar1-1、jar1-2和jar1-4经土壤腐霉菌侵染后, 病症比野生型更明显, 并表现对外源JA的敏感性降低及内源JA-Ile水平降低, 说明JAR1参与JA信号转导途径, 增强植物抗病性。对JAR1的进一步研究表明, JAR1参与非致病性根瘤菌引发的系统获得抗性(systemic acquired resistance, SAR), 还能减少臭氧对拟南芥的损伤(van Loon et al., 1998; Overmyer et al., 2000; Rao et al., 2000)。Zhang等(2007)发现AtGH3.5除调节IAA含量外, 还参与SA信号转导途径, AtGH3.5过表达导致SA积累增加, 及响应无毒病原体的病程相关蛋白PR-1表达量增加。研究表明, atgh3.5通过依赖NPR1和独立途径增强SA介导的防御反应(Zhang et al., 2008)。此外, 在AtGH3.12/PBS3等位基因GDG1突变体gdg1中, SA依赖性基因表达和防御受损, 响应病原体的SA积累受抑制。gdg1所有防御相关表型都可通过外源施加SA来挽救, 表明GDG1位于SA上游, 调节SA水平, 进而增强对病原体的抗性(Jagadeeswaran et al., 2007; Okrent et al., 2009)。AtGH3.6过表达株系游离IAA的含量减少, 导致抗旱性和耐盐性均低于野生型; 在干旱、ABA和高盐胁迫下, AtGH3.6基因的高表达抑制逆境响应基因RD22、KIN1、RD29A和DREB1A的表达, 表明AtGH3.6负调控拟南芥逆境响应(刘晓东等, 2016)。光是影响植物生长发育的重要环境因素之一。植物响应外界光信号, 通过合成生长素等植物激素进一步灵活地调控基因表达。外源光信号和生长素信号通路之间的整合使植物得以协调外界环境刺激与内在生长发育, 达到适应环境、优化生长的目的。Xu等(2018)发现了生长素信号与光信号互作的新机制——蓝光受体CRY1和光敏色素B (phyB)分别与AUX/IAA互作, 抑制生长素引起的AUX/IAA蛋白降解, 减弱生长素信号, 进而抑制下胚轴伸长。拟南芥GH3基因突变后, 在受到光信号调控时表现出一定的表型。dfl1-D在蓝光、红光和远红光下具有较短的下胚轴, 且侧根生长受抑制(Nakazawa et al., 2001)。红光和蓝光下DFL2过表达株系同样表现出短的下胚轴, 而在红光下反义表达时导致长下胚轴表型(Takase et al., 2003)。远红光下AtGH3.5过表达株系wes1-D下胚轴明显比对照短, 但突变体wes1下胚轴比对照长(Park et al., 2007b)。综上, AtGH3s基因参与生长素与光信号途径, 共同调节拟南芥下胚轴伸长。此外, AtGH3.11等位基因FIN219突变体fin219在连续远红光下表现长下胚轴表型, 并作用于光形态建成抑制因子COP1的上游, 推测远红光通过调控FIN219基因, 进而负调控COP1 (McNellis et al., 1994; Osterlund et al., 1999; Hsieh et al., 2000)。
水稻株型是决定其产量的核心因素之一。水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等。其中, 生长素的含量和分布对植株形态建成产生重要影响。水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量。OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008)。TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少。在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009)。OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012)。Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根。此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013)。
病原菌感染可诱导IAA积累, 而IAA诱导膨胀蛋白的表达, 使细胞壁松弛。松弛的细胞壁是植物生长的关键, 但也使植物容易受到病原菌感染。OsGH3.1和OsGH3.8过表达引起游离IAA含量显著降低, 抑制细胞生长和细胞壁松弛, 从而增强水稻对病原菌的抗性(Ding et al., 2008; Domingo et al., 2009)。OsGH3.2过表达株系游离IAA含量降低, 氧化损伤减轻, 质膜透性降低, 表现出较强的耐冷性, 并对干旱胁迫敏感(Du et al., 2012)。OsWRKY13和OsWRKY45是JA信号响应正调控基因(Qiu et al., 2007; Tao et al., 2009)。Hui等(2019)发现这2个基因在OsGH3.3、OsGH3.5、OsGH3.6和OsGH3.12过表达株系中被显著激活表达, 表明这4个基因通过调节JA响应基因的表达增强水稻对白叶枯病菌的抗性。OsGH3.5/ OsJAR1为AtJAR1的同源基因, 在水稻JA和光信号转导途径中发挥相同作用(Riemann et al., 2008)。在远红光和蓝光下, osjar1突变体胚芽鞘比野生型更长, 表明OsJAR1参与光下胚芽鞘伸长的抑制。
在茄科植物番茄中鉴定到15个GH3家族基因.王慧敏(2015)研究发现, 与野生型相比, SlGH3.2过表达株系明显矮化, 结实率低, 且种子显著变小.SlGH3.4过表达株系根中游离IAA浓度降低, 显著抑制菌根共生和一些生长素响应基因表达, 同时丛枝表现为生长素缺陷的发育畸形(陈潇, 2017).在番茄中过表达SlGH3.15基因, 过表达株系表现出由节间距缩短所致的矮化、叶片卷曲及侧根变少, 且过表达植株体内游离IAA含量减少(艾国, 2017).番茄YABBY2b转录因子直接结合SlGH3.8启动子并抑制其表达, 正调控植物生长; yabby2b突变体表现出矮化和较小的花和果实, 表明SlGH3.8负调控植物的生长发育(Sun et al., 2020). ...
在茄科植物番茄中鉴定到15个GH3家族基因.王慧敏(2015)研究发现, 与野生型相比, SlGH3.2过表达株系明显矮化, 结实率低, 且种子显著变小.SlGH3.4过表达株系根中游离IAA浓度降低, 显著抑制菌根共生和一些生长素响应基因表达, 同时丛枝表现为生长素缺陷的发育畸形(陈潇, 2017).在番茄中过表达SlGH3.15基因, 过表达株系表现出由节间距缩短所致的矮化、叶片卷曲及侧根变少, 且过表达植株体内游离IAA含量减少(艾国, 2017).番茄YABBY2b转录因子直接结合SlGH3.8启动子并抑制其表达, 正调控植物生长; yabby2b突变体表现出矮化和较小的花和果实, 表明SlGH3.8负调控植物的生长发育(Sun et al., 2020). ...
在茄科植物番茄中鉴定到15个GH3家族基因.王慧敏(2015)研究发现, 与野生型相比, SlGH3.2过表达株系明显矮化, 结实率低, 且种子显著变小.SlGH3.4过表达株系根中游离IAA浓度降低, 显著抑制菌根共生和一些生长素响应基因表达, 同时丛枝表现为生长素缺陷的发育畸形(陈潇, 2017).在番茄中过表达SlGH3.15基因, 过表达株系表现出由节间距缩短所致的矮化、叶片卷曲及侧根变少, 且过表达植株体内游离IAA含量减少(艾国, 2017).番茄YABBY2b转录因子直接结合SlGH3.8启动子并抑制其表达, 正调控植物生长; yabby2b突变体表现出矮化和较小的花和果实, 表明SlGH3.8负调控植物的生长发育(Sun et al., 2020). ...
在茄科植物番茄中鉴定到15个GH3家族基因.王慧敏(2015)研究发现, 与野生型相比, SlGH3.2过表达株系明显矮化, 结实率低, 且种子显著变小.SlGH3.4过表达株系根中游离IAA浓度降低, 显著抑制菌根共生和一些生长素响应基因表达, 同时丛枝表现为生长素缺陷的发育畸形(陈潇, 2017).在番茄中过表达SlGH3.15基因, 过表达株系表现出由节间距缩短所致的矮化、叶片卷曲及侧根变少, 且过表达植株体内游离IAA含量减少(艾国, 2017).番茄YABBY2b转录因子直接结合SlGH3.8启动子并抑制其表达, 正调控植物生长; yabby2b突变体表现出矮化和较小的花和果实, 表明SlGH3.8负调控植物的生长发育(Sun et al., 2020). ...
在茄科植物番茄中鉴定到15个GH3家族基因.王慧敏(2015)研究发现, 与野生型相比, SlGH3.2过表达株系明显矮化, 结实率低, 且种子显著变小.SlGH3.4过表达株系根中游离IAA浓度降低, 显著抑制菌根共生和一些生长素响应基因表达, 同时丛枝表现为生长素缺陷的发育畸形(陈潇, 2017).在番茄中过表达SlGH3.15基因, 过表达株系表现出由节间距缩短所致的矮化、叶片卷曲及侧根变少, 且过表达植株体内游离IAA含量减少(艾国, 2017).番茄YABBY2b转录因子直接结合SlGH3.8启动子并抑制其表达, 正调控植物生长; yabby2b突变体表现出矮化和较小的花和果实, 表明SlGH3.8负调控植物的生长发育(Sun et al., 2020). ...
在茄科植物番茄中鉴定到15个GH3家族基因.王慧敏(2015)研究发现, 与野生型相比, SlGH3.2过表达株系明显矮化, 结实率低, 且种子显著变小.SlGH3.4过表达株系根中游离IAA浓度降低, 显著抑制菌根共生和一些生长素响应基因表达, 同时丛枝表现为生长素缺陷的发育畸形(陈潇, 2017).在番茄中过表达SlGH3.15基因, 过表达株系表现出由节间距缩短所致的矮化、叶片卷曲及侧根变少, 且过表达植株体内游离IAA含量减少(艾国, 2017).番茄YABBY2b转录因子直接结合SlGH3.8启动子并抑制其表达, 正调控植物生长; yabby2b突变体表现出矮化和较小的花和果实, 表明SlGH3.8负调控植物的生长发育(Sun et al., 2020). ...
在茄科植物番茄中鉴定到15个GH3家族基因.王慧敏(2015)研究发现, 与野生型相比, SlGH3.2过表达株系明显矮化, 结实率低, 且种子显著变小.SlGH3.4过表达株系根中游离IAA浓度降低, 显著抑制菌根共生和一些生长素响应基因表达, 同时丛枝表现为生长素缺陷的发育畸形(陈潇, 2017).在番茄中过表达SlGH3.15基因, 过表达株系表现出由节间距缩短所致的矮化、叶片卷曲及侧根变少, 且过表达植株体内游离IAA含量减少(艾国, 2017).番茄YABBY2b转录因子直接结合SlGH3.8启动子并抑制其表达, 正调控植物生长; yabby2b突变体表现出矮化和较小的花和果实, 表明SlGH3.8负调控植物的生长发育(Sun et al., 2020). ...
在茄科植物番茄中鉴定到15个GH3家族基因.王慧敏(2015)研究发现, 与野生型相比, SlGH3.2过表达株系明显矮化, 结实率低, 且种子显著变小.SlGH3.4过表达株系根中游离IAA浓度降低, 显著抑制菌根共生和一些生长素响应基因表达, 同时丛枝表现为生长素缺陷的发育畸形(陈潇, 2017).在番茄中过表达SlGH3.15基因, 过表达株系表现出由节间距缩短所致的矮化、叶片卷曲及侧根变少, 且过表达植株体内游离IAA含量减少(艾国, 2017).番茄YABBY2b转录因子直接结合SlGH3.8启动子并抑制其表达, 正调控植物生长; yabby2b突变体表现出矮化和较小的花和果实, 表明SlGH3.8负调控植物的生长发育(Sun et al., 2020). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
在茄科植物番茄中鉴定到15个GH3家族基因.王慧敏(2015)研究发现, 与野生型相比, SlGH3.2过表达株系明显矮化, 结实率低, 且种子显著变小.SlGH3.4过表达株系根中游离IAA浓度降低, 显著抑制菌根共生和一些生长素响应基因表达, 同时丛枝表现为生长素缺陷的发育畸形(陈潇, 2017).在番茄中过表达SlGH3.15基因, 过表达株系表现出由节间距缩短所致的矮化、叶片卷曲及侧根变少, 且过表达植株体内游离IAA含量减少(艾国, 2017).番茄YABBY2b转录因子直接结合SlGH3.8启动子并抑制其表达, 正调控植物生长; yabby2b突变体表现出矮化和较小的花和果实, 表明SlGH3.8负调控植物的生长发育(Sun et al., 2020). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
在茄科植物番茄中鉴定到15个GH3家族基因.王慧敏(2015)研究发现, 与野生型相比, SlGH3.2过表达株系明显矮化, 结实率低, 且种子显著变小.SlGH3.4过表达株系根中游离IAA浓度降低, 显著抑制菌根共生和一些生长素响应基因表达, 同时丛枝表现为生长素缺陷的发育畸形(陈潇, 2017).在番茄中过表达SlGH3.15基因, 过表达株系表现出由节间距缩短所致的矮化、叶片卷曲及侧根变少, 且过表达植株体内游离IAA含量减少(艾国, 2017).番茄YABBY2b转录因子直接结合SlGH3.8启动子并抑制其表达, 正调控植物生长; yabby2b突变体表现出矮化和较小的花和果实, 表明SlGH3.8负调控植物的生长发育(Sun et al., 2020). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
(A) Under higher IAA concentration, ARF splits with Aux/IAAs dimer, and ARF binds to AuxREs and activates the expression of GH3, which catalyzes the binding of IAA to amino acids; (B) Under lower IAA concentration, ARF forms a dimer with Aux/IAAs and turns off AuxREs, GH3 gene transcription is inhibited, and IAA-Ala and IAA-Leu, which are auxin reservoirs, are hydrolyzed by amidohydrolase to release IAA again. ... Boosting CRISPR/ Cas9 multiplex editing capability with the endogenous tRNA-processing system 0 2015 Photoactivated CRY1 and phyB interact directly with AUX/IAA proteins to inhibit auxin signaling in Arabidopsis 1 2018 ... 植物在生长发育过程中常受到生物与非生物胁迫, JA和SA作为信号分子在植物胁迫响应中发挥重要作用.目前, GH3基因参与拟南芥胁迫响应的研究主要集中在少数几个AtGH3s基因中.Staswick等(1998)研究发现, JAR1/AtGH3.11突变体jar1-1、jar1-2和jar1-4经土壤腐霉菌侵染后, 病症比野生型更明显, 并表现对外源JA的敏感性降低及内源JA-Ile水平降低, 说明JAR1参与JA信号转导途径, 增强植物抗病性.对JAR1的进一步研究表明, JAR1参与非致病性根瘤菌引发的系统获得抗性(systemic acquired resistance, SAR), 还能减少臭氧对拟南芥的损伤(van Loon et al., 1998; Overmyer et al., 2000; Rao et al., 2000).Zhang等(2007)发现AtGH3.5除调节IAA含量外, 还参与SA信号转导途径, AtGH3.5过表达导致SA积累增加, 及响应无毒病原体的病程相关蛋白PR-1表达量增加.研究表明, atgh3.5通过依赖NPR1和独立途径增强SA介导的防御反应(Zhang et al., 2008).此外, 在AtGH3.12/PBS3等位基因GDG1突变体gdg1中, SA依赖性基因表达和防御受损, 响应病原体的SA积累受抑制.gdg1所有防御相关表型都可通过外源施加SA来挽救, 表明GDG1位于SA上游, 调节SA水平, 进而增强对病原体的抗性(Jagadeeswaran et al., 2007; Okrent et al., 2009).AtGH3.6过表达株系游离IAA的含量减少, 导致抗旱性和耐盐性均低于野生型; 在干旱、ABA和高盐胁迫下, AtGH3.6基因的高表达抑制逆境响应基因RD22、KIN1、RD29A和DREB1A的表达, 表明AtGH3.6负调控拟南芥逆境响应(刘晓东等, 2016).光是影响植物生长发育的重要环境因素之一.植物响应外界光信号, 通过合成生长素等植物激素进一步灵活地调控基因表达.外源光信号和生长素信号通路之间的整合使植物得以协调外界环境刺激与内在生长发育, 达到适应环境、优化生长的目的.Xu等(2018)发现了生长素信号与光信号互作的新机制——蓝光受体CRY1和光敏色素B (phyB)分别与AUX/IAA互作, 抑制生长素引起的AUX/IAA蛋白降解, 减弱生长素信号, 进而抑制下胚轴伸长.拟南芥GH3基因突变后, 在受到光信号调控时表现出一定的表型.dfl1-D在蓝光、红光和远红光下具有较短的下胚轴, 且侧根生长受抑制(Nakazawa et al., 2001).红光和蓝光下DFL2过表达株系同样表现出短的下胚轴, 而在红光下反义表达时导致长下胚轴表型(Takase et al., 2003).远红光下AtGH3.5过表达株系wes1-D下胚轴明显比对照短, 但突变体wes1下胚轴比对照长(Park et al., 2007b).综上, AtGH3s基因参与生长素与光信号途径, 共同调节拟南芥下胚轴伸长.此外, AtGH3.11等位基因FIN219突变体fin219在连续远红光下表现长下胚轴表型, 并作用于光形态建成抑制因子COP1的上游, 推测远红光通过调控FIN219基因, 进而负调控COP1 (McNellis et al., 1994; Osterlund et al., 1999; Hsieh et al., 2000). ... Dynamic perception of jasmonates by the F-box protein COI1 1 2018 ... GH3亚家族I催化JA与氨基酸结合, 形成具有生物活性的茉莉酸异亮氨酸复合物(JA-Ile) (Staswick et al., 2002).茉莉酸是植物体内重要的脂质激素, 在植物胁迫响应中发挥重要作用.同时, 茉莉酸作为重要的生长发育调节物质, 参与调节植物生长发育的诸多过程(Wasternack and Hause, 2013; 李梦莎和阎秀峰, 2014; 黎家和李传友, 2019).当植物受到机械损伤或昆虫啃食时, 受伤部位迅速积累的JA被JAR1/AtGH3.11催化形成JA信号转导中的主要活性复合物JA-Ile.JA-Ile先被SCFCOI1受体识别, 后招募茉莉酸响应抑制因子JAZ (jasmonate ZIM-domain), 形成SCFCOI1-JA-Ile-JAZ三元复合物(Yan et al., 2018).形成的三元复合物使JAZ被26S蛋白降解, 从而解除对转录因子(transcription factors, TF) MYC2的抑制作用, 激活JA响应基因的表达(图1) (李梦莎和阎秀峰, 2014). ... Evidence of an auxin signal pathway, microRNA167-ARF8-GH3, and its response to exogenous auxin in cultured rice cells 3 2006 ... Functions of GH3 genes in rice
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
水稻株型是决定其产量的核心因素之一.水稻株型的形成主要取决于植株高度、分蘖数及分蘖角度等.其中, 生长素的含量和分布对植株形态建成产生重要影响.水稻根系类型为须根系, 不定根和侧根是其生长后期营养和水分吸收的主要器官, 根系发育同样影响水稻生长发育及产量.OsGH3.8过表达植株表现出矮化和分蘖数增加, 且具有更短的根和更少的不定根, 可育性和种子发芽率大幅降低(Ding et al., 2008).TLD1/OsGH3.13功能获得突变体tld1-D分蘖数增加, 叶夹角增大, 矮化, 还表现出细秆、种子细长、短穗和侧根减少.在正常生长条件下, 地上部组织中TLD1被抑制, 但在干旱胁迫下被显著诱导(Zhang et al., 2009).OsGH3.2过表达株系表现出矮化、叶片变小、不定根和根毛减少等表型(Du et al., 2012).Liu等(2022)发现, 与野生型相比, osgh3.2突变体表现出短根, 且具有更短的不定根及更多的侧根与不定根.此外, 水稻中过表达OsGH3.1获得LC1过表达株系, 因其游离IAA含量减少而表现出半矮化和叶夹角增大等表型, 且对BR信号敏感(Zhao et al., 2013). ...
本文的其它图/表
-
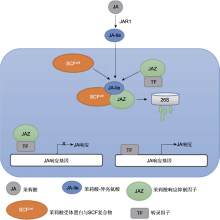 图1
茉莉酸信号转导通路(参考李梦莎和阎秀峰, 2014; Lee et al., 2022)
图1
茉莉酸信号转导通路(参考李梦莎和阎秀峰, 2014; Lee et al., 2022)
-
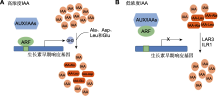 图2
植物体内生长素(IAA)动态平衡(参考Hagen and Guilfoyle, 2002; Woodward and Bartel, 2005)
图2
植物体内生长素(IAA)动态平衡(参考Hagen and Guilfoyle, 2002; Woodward and Bartel, 2005)
(A) IAA浓度较高时, ARF与Aux/IAAs二聚体分离, ARF结合到相应的AuxREs并激活GH3表达, 催化IAA与氨基酸结合; (B) IAA浓度较低时, ARF与Aux/IAAs形成二聚体并关闭AuxREs, GH3基因转录受到抑制, 作为生长素贮存库的IAA-Ala和IAA-Leu由酰胺水解酶水解, 重新释放出IAA。
-
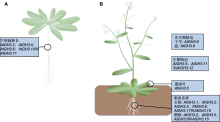 图3
拟南芥生长发育中GH3基因的功能(参考Staswick et al., 1998, 2005; van Loon et al., 1998; Hsieh et al., 2000; Overmyer et al., 2000; Rao et al., 2000; Nakazawa et al., 2001; Takase et al., 2003, 2004; Park et al., 2007b; Jagadeeswaran et al., 2007; Nobuta et al., 2007; Zhang et al., 2007; 2008; 周苹等, 2015; 刘晓东等, 2016; Sherp et al., 2018; 周淑瑶等, 2023)
图3
拟南芥生长发育中GH3基因的功能(参考Staswick et al., 1998, 2005; van Loon et al., 1998; Hsieh et al., 2000; Overmyer et al., 2000; Rao et al., 2000; Nakazawa et al., 2001; Takase et al., 2003, 2004; Park et al., 2007b; Jagadeeswaran et al., 2007; Nobuta et al., 2007; Zhang et al., 2007; 2008; 周苹等, 2015; 刘晓东等, 2016; Sherp et al., 2018; 周淑瑶等, 2023)
(A) 拟南芥幼苗; (B) 拟南芥成熟植株
-
 表2
水稻GH3基因的生物学功能
表2
水稻GH3基因的生物学功能
-
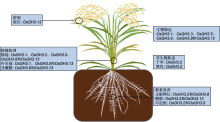 图4
水稻生长发育中GH3基因的功能(参考Yang et al., 2006; Qiu et al., 2007; Ding et al., 2008; Domingo et al., 2009; Tao et al., 2009; Zhang et al., 2009, 2015; Du et al., 2012; Zhao et al., 2013; Dai et al., 2018; Hui et al., 2019; Liu et al., 2022)
图4
水稻生长发育中GH3基因的功能(参考Yang et al., 2006; Qiu et al., 2007; Ding et al., 2008; Domingo et al., 2009; Tao et al., 2009; Zhang et al., 2009, 2015; Du et al., 2012; Zhao et al., 2013; Dai et al., 2018; Hui et al., 2019; Liu et al., 2022)
-
表3
其它植物GH3基因的生物学功能
|

 首页
首页