
|
|
||
|
植物GH3基因家族生物学功能研究进展
植物学报
2023, 58 (5):
770-782.
DOI: 10.11983/CBB22263
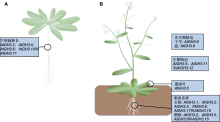
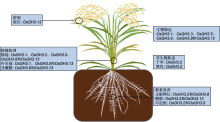
植物生长素早期响应基因GH3编码的酰胺合酶催化生长素、茉莉酸及苯甲酸衍生物与氨基酸结合, 形成相应的氨基酸复合物。当植物体内生长素浓度过高时, GH3蛋白催化生长素与氨基酸结合, 形成的复合物作为生长素贮存库。当生长素浓度过低时, 生长素-氨基酸复合物被蛋白水解酶水解为生长素, 重新参与生长素信号通路, 从而调控植物体内生长素动态平衡。当植物受到生物或非生物胁迫时, GH3蛋白催化茉莉酸和水杨酸与氨基酸结合, 参与植物胁迫响应。该文从GH3蛋白结构、GH3基因家族分类及其生物学功能方面总结了双子叶模式植物拟南芥(Arabidopsis thaliana)、单子叶模式植物水稻(Oryza sativa)及其它植物中GH3基因的研究进展, 为植物GH3基因家族的深入研究提供参考。
 View image in article
图1
茉莉酸信号转导通路(参考李梦莎和阎秀峰,
正文中引用本图/表的段落
GH3亚家族I催化JA与氨基酸结合, 形成具有生物活性的茉莉酸异亮氨酸复合物(JA-Ile) (Staswick et al., 2002)。茉莉酸是植物体内重要的脂质激素, 在植物胁迫响应中发挥重要作用。同时, 茉莉酸作为重要的生长发育调节物质, 参与调节植物生长发育的诸多过程(Wasternack and Hause, 2013; 李梦莎和阎秀峰, 2014; 黎家和李传友, 2019)。当植物受到机械损伤或昆虫啃食时, 受伤部位迅速积累的JA被JAR1/AtGH3.11催化形成JA信号转导中的主要活性复合物JA-Ile。JA-Ile先被SCFCOI1受体识别, 后招募茉莉酸响应抑制因子JAZ (jasmonate ZIM-domain), 形成SCFCOI1-JA-Ile-JAZ三元复合物(Yan et al., 2018)。形成的三元复合物使JAZ被26S蛋白降解, 从而解除对转录因子(transcription factors, TF) MYC2的抑制作用, 激活JA响应基因的表达(图1) (李梦莎和阎秀峰, 2014)。
(A) Under higher IAA concentration, ARF splits with Aux/IAAs dimer, and ARF binds to AuxREs and activates the expression of GH3, which catalyzes the binding of IAA to amino acids; (B) Under lower IAA concentration, ARF forms a dimer with Aux/IAAs and turns off AuxREs, GH3 gene transcription is inhibited, and IAA-Ala and IAA-Leu, which are auxin reservoirs, are hydrolyzed by amidohydrolase to release IAA again. ... Boosting CRISPR/ Cas9 multiplex editing capability with the endogenous tRNA-processing system 0 2015 Photoactivated CRY1 and phyB interact directly with AUX/IAA proteins to inhibit auxin signaling in Arabidopsis 1 2018 ... 植物在生长发育过程中常受到生物与非生物胁迫, JA和SA作为信号分子在植物胁迫响应中发挥重要作用.目前, GH3基因参与拟南芥胁迫响应的研究主要集中在少数几个AtGH3s基因中.Staswick等( Dynamic perception of jasmonates by the F-box protein COI1 1 2018 ... GH3亚家族I催化JA与氨基酸结合, 形成具有生物活性的茉莉酸异亮氨酸复合物(JA-Ile) (Staswick et al., Evidence of an auxin signal pathway, microRNA167-ARF8-GH3, and its response to exogenous auxin in cultured rice cells 3 2006 ... Functions of GH3 genes in rice
本文的其它图/表
|
 首页
首页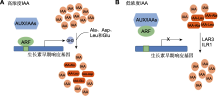
{kind=link}