|
|
|||||||||||||||||||||||||||||
|
山西小麦籽粒叶黄素含量变异及遗传特性分析
植物学报
2023, 58 (4):
535-547.
DOI: 10.11983/CBB22081
叶黄素在保护视力、预防糖尿病、心血管硬化和癌症等方面具有良好功效。利用小麦(Triticum aestivum)进行叶黄素的生物强化逐渐引起人们的重视。以3种环境下种植的194份山西小麦为材料, 采用有机溶剂浸提法提取叶黄素, 利用高效液相色谱法测定不同种质的叶黄素含量, 分析小麦叶黄素含量的广义遗传力, 及其与籽粒颜色、冬春性、地域分布、品种类型和主要农艺性状的关系, 并通过全基因组关联分析挖掘叶黄素含量相关的遗传位点。结果表明, 山西小麦品种间叶黄素含量变异范围较广, 变异系数为33.12%-48.57%, 基因型是影响叶黄素含量的主要因素, 3种环境下小麦叶黄素含量范围分别为0.67-4.03、0.16-5.05和0.16-3.63 μg·g-1; 冬性小麦品种的平均含量高于春性品种, 水地品种平均含量高于旱地品种, 籽粒颜色与育种年代对叶黄素含量无显著影响; 抽穗期、株高和千粒重与叶黄素含量呈显著负相关, 其它农艺性状对叶黄素均无明显影响; 全基因组关联分析在1B、3A和7A染色体上发现4个与叶黄素含量相关的主效位点, 其中QLuc.3A和QLuc.7A.1是影响叶黄素含量的新位点。研究结果可为小麦叶黄素生物强化品种的选育和栽培提供有价值的信息。
表3
不同环境下山西小麦叶黄素含量分组
正文中引用本图/表的段落
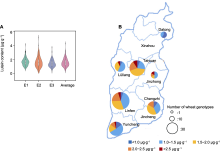
将3种环境下收获的材料提取叶黄素进行测定, 发现山西小麦品种间的叶黄素含量变异范围较广(附表1), 变异系数为33.84%-43.90%; 含量范围分别为0.67- 4.03、0.16-5.05和0.16-3.63 μg·g-1, 平均值分别为1.63、1.57和1.39 μg·g-1 (图2A)。根据叶黄素含量将供试材料分为5级(<1.0 (μg·g-1)、1.0-1.5 (μg·g-1)、1.5-2.0 (μg·g-1)、2.0-2.5 (μg·g-1)和>2.5 (μg·g-1)), 其中含量在1.0-1.5和1.5-2.0 μg·g-1范围内的品种较多, 占总品种数的62%, 超过2.5 μg·g-1的品种数分别为11、24和3份(表3)。E1环境下含量最高的3个品种分别为河东TX-006、长4853和晋麦95号(图1C); E2环境下为河东TX-006、太113和山农129; E3环境下为河东TX-006、晋麦95号和临远3158。其中, 河东TX-006、晋麦95号和临远3158在3种环境下的叶黄素含量均超过3.0 μg·g-1, 是高叶黄素含量品种。
(A) 3种环境下小麦品种的叶黄素含量分布; (B) 不同地域品种的叶黄素含量分布。E1、E2和E3同
研究基因型、环境及其互作对3种环境下叶黄素含量的影响, 结果表明, 叶黄素含量广义遗传力在基因型效应(G)、环境效应(E)和基因型与环境互作效应(G×E)均达到极显著水平。基因型效应对叶黄素含量的影响最大, 籽粒叶黄素含量主要受基因型的影响, 其次为环境; 3个因素具体表现为基因型效应(67.58%) >环境效应(31.93%) >基因型与环境互作效应(0.49%)。
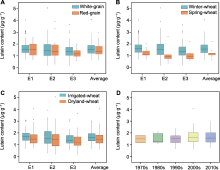
根据籽粒颜色将种质分为红粒和白粒, 比较后发现不同粒色品种间叶黄素含量差异不显著, 除E1环境中红粒品种叶黄素含量略高于白粒外, 其余环境均是白粒品种稍高于红粒(图3A)。小麦主要为冬性和春性品种2种类型, 比较发现冬性品种叶黄素含量均显著高于春性品种, 其中E1和E2环境中冬、春性间叶黄素含量差异极显著(图3B), 可能是冬性品种生育周期长, 使得籽粒中积累了较多的叶黄素。山西小麦品种类型主要分为旱地和水地两种, 不同环境下水地品种叶黄素含量均高于旱地品种, 其中E1和E2环境下水旱品种的含量差异显著(图3C), 原因可能是水地品种普遍较旱地品种成熟期晚, 灌浆期长有利于叶黄素的积累, 这与冬性品种高于春性品种的结果一致。分析不同年代育成品种叶黄素含量, 发现叶黄素含量在选育过程中无明显变化, 表明叶黄素在山西小麦育种历程中并未受到人工和自然选择(图3D)。
E1、E2和E3同
E1、E2和E3同
E1、E2和E3同
E1、E2和E3同
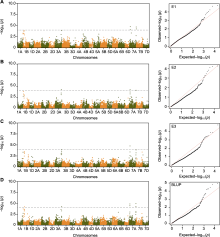
多年以来, 小麦籽粒叶黄素含量的遗传研究主要集中在四倍体小麦, 对六倍体小麦研究较少, 且主要以黄色素混合物为研究对象。已有研究采用SSR和AFLP标记及90K、660K SNP芯片, 利用硬粒及普通小麦的不同遗传和自然群体, 将影响黄色素的QTL定位在除1D、4D和6D外的染色体上(Elouafi et al., 2001; Pozniak et al., 2007; Patil et al., 2008; Zhang et al., 2008; Reimer et al., 2008; Singh et al., 2009; Blanco et al., 2011; Zhao et al., 2013; Colasuonno et al., 2014, 2017, 2019; Zhai et al., 2016)。其中, 第7同源群上的QTL可解释50%以上的表型变异, 3B和5B上的QTL可解释12%-20%的表型变异(Mares and Campbell, 2001; Blanco et al., 2011)。此外, ε-环化酶(ε-LCY1)和八氢番茄红素合成酶(Psy-A1)的小麦同源基因中, ε-LCY的点突变导致1个保守氨基酸替换, 引起叶黄素含量升高; Psy-A1可变剪接导致编码蛋白提前终止或者移码突变使蛋白失活, 继而影响籽粒叶黄素的积累(Howitt et al., 2009; Dibari et al., 2012; Crawford and Francki, 2013)。叶黄素属于次生代谢产物, 参与植物的生长发育、逆境响应和损伤修复等多种生物学过程, 其合成和降解受许多途径的调节(Demmig-Adams and Adams, 2002; Li et al., 2008; Welsch et al., 2008; Farré et al., 2010)。本研究利用GWAS检测到4个影响小麦叶黄素含量的QTL位点, QLuc.7A.2和QLuc.1B是已报道的位点(Howitt et al., 2009; Li et al., 2016), QLuc.3A和QLuc.7A.1是2个新的叶黄素含量相关位点, 可用于开发分子标记, 为今后小麦叶黄素生物强化品种的选育奠定基础。
本文的其它图/表
|
|||||||||||||||||||||||||||||
 首页
首页