
|
|
||
|
植物细胞壁木聚糖的生物合成及其应用
植物学报
2023, 58 (2):
316-334.
DOI: 10.11983/CBB22030
木聚糖是广泛存在于各类植物细胞壁中的半纤维素, 对植物生长发育至关重要。许多研究表明, 细胞壁中木聚糖的含量和结构对生物质的加工特性有显著影响。因此, 理解木聚糖的生物合成机制有助于利用基因工程手段对细胞壁进行改良。近10年来, 在模式植物拟南芥(Arabidopsis thaliana)以及重要粮食和经济作物中鉴定出许多参与木聚糖生物合成的基因。该文综述了相关研究进展, 并探讨了木聚糖生物合成基因在生物质能及相关领域的潜在应用价值。 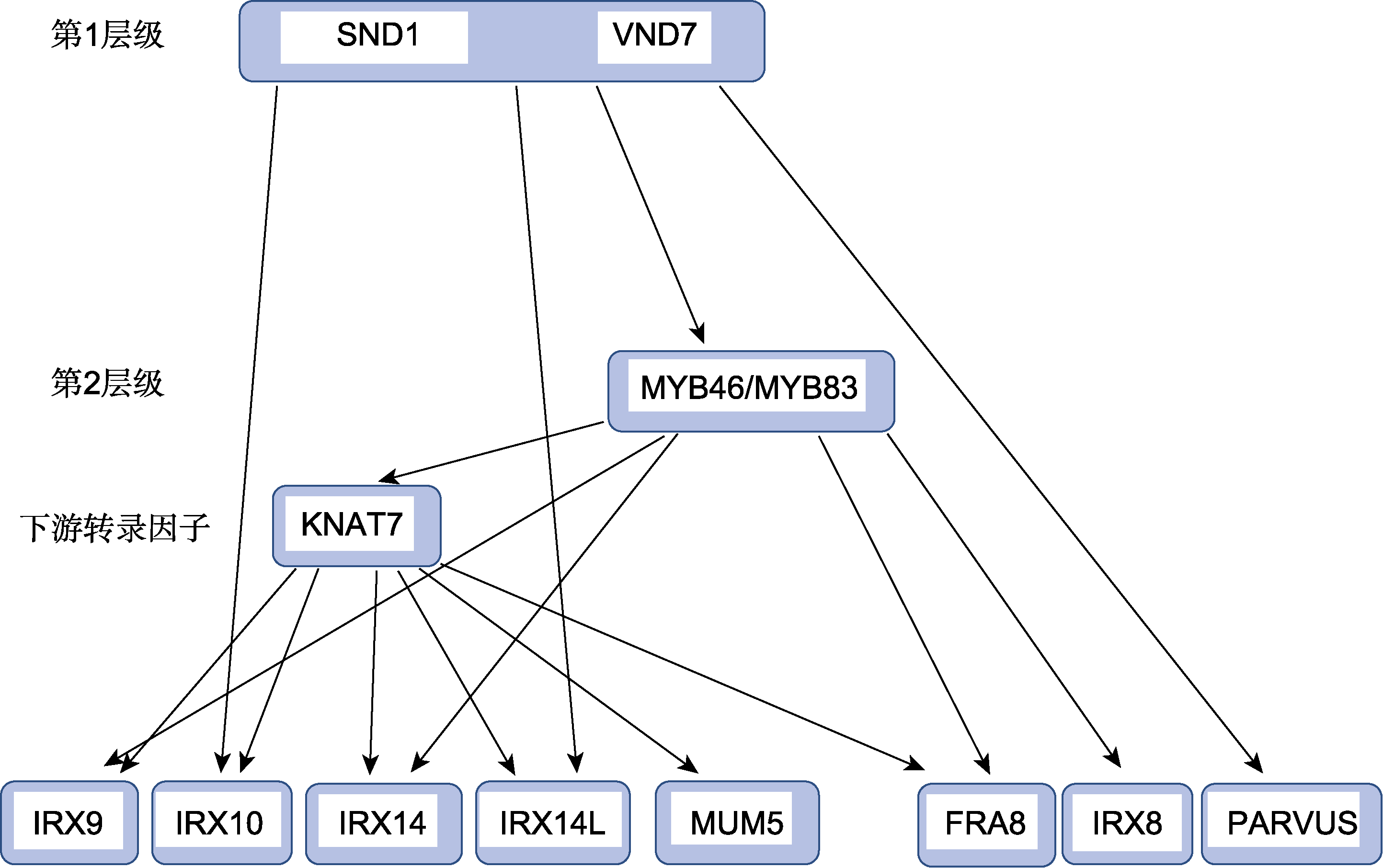 View image in article
图3
木聚糖生物合成的转录调控示意图
正文中引用本图/表的段落
植物次生壁的生物合成由NAC及MYB等转录因子组成的层级网络所调控。第1层级是NAC类转录因子, 如NST1、SND1/NST3、VND6和VND7; 第2层级是MYB类转录因子MYB46/MYB83, 它们可以直接或间接地调控次生壁的合成, 是主开关转录因子, 在其下游还有一些转录因子, 特异性地调控某些基因的表达(Nakano et al., 2015; 张雨等, 2020)。木聚糖作为次生壁半纤维素的主要成分, 其合成受该层级网络中的转录因子调控。已有证据表明, 位于第1层级的SND1和VND7、第1层级的直接靶标MYB46/MYB83以及MYB46的直接靶标KNAT7可以调控拟南芥木聚糖的合成(图3)。
ND: 尚未确定. ... NAC-MYB-based transcriptional regulation of secondary cell wall biosynthesis in land plants 1 2015 ... 植物次生壁的生物合成由NAC及MYB等转录因子组成的层级网络所调控.第1层级是NAC类转录因子, 如NST1、SND1/NST3、VND6和VND7; 第2层级是MYB类转录因子MYB46/MYB83, 它们可以直接或间接地调控次生壁的合成, 是主开关转录因子, 在其下游还有一些转录因子, 特异性地调控某些基因的表达(Nakano et al., Designing xylan for improved sustainable biofuel production 1 2019 ... 木聚糖是细胞壁中主要的半纤维素, 可以通过氢键与纤维素结合, 通过氢键和共价键与木质素和果胶结合, 参与细胞壁的形成, 决定细胞壁的物理性质(Tan et al., In muro deacetylation of xylan affects lignin properties and improves saccharification of aspen wood 1 2017 ... 将从黑曲霉(Aspergillus niger)中分离出的木聚糖乙酰酯酶基因AnAXE1转入杂种白杨(P. tremula × tremuloides), 可适度降低杂种白杨细胞壁中木聚糖的乙酰化程度, 使糖化作用得到增强而植物生长不受影响(Pawar et al., Acetylation of woody lignocellulose: significance and regulation 1 2013 ... 硬木木聚糖主链的C-2和/或C-3位置有70%会发生乙酰化, 而软木木聚糖主链通常缺乏乙酰化(Pawar et al., Cell walls of developing wheat starchy endosperm: comparison of composition and RNA-seq transcriptome 1 2012 ... 小麦胚乳细胞壁的主要成分是阿拉伯木聚糖, 约占细胞壁总多糖的70%.小麦籽粒水提物(water-extractable, WE-AX)是饮食结构中可溶性膳食纤维的来源, 也是酿酒工业的重要原料.WE-AX黏度是影响各种加工过程的关键性状, 其很大程度取决于胚乳AX的性质.在小麦胚乳中表达量最高的IRX14、IRX9和IRX10的同源基因分别为TaGT43_1、TaGT43_2和TaGT47_2, 它们是胚乳中最有可能参与木聚糖主链合成的基因(Pellny et al., Loss of TaIRX9b gene function in wheat decreases chain length and amount of arabinoxylan in grain but increases cross-linking 4 2020 ... 木聚糖的主链绝大多数是由木糖(xylose, Xyl)通过β-1,4糖苷键连接成的糖链(Pe?a et al.,
本文的其它图/表
|
 首页
首页
{kind=link}