植物细胞壁木聚糖的生物合成及其应用
郭彦君, 陈枫, 罗敬文, 曾为, 许文亮
植物学报
2023, 58 ( 2):
316-334.
DOI: 10.11983/CBB22030
木聚糖是广泛存在于各类植物细胞壁中的半纤维素, 对植物生长发育至关重要。许多研究表明, 细胞壁中木聚糖的含量和结构对生物质的加工特性有显著影响。因此, 理解木聚糖的生物合成机制有助于利用基因工程手段对细胞壁进行改良。近10年来, 在模式植物拟南芥(Arabidopsis thaliana)以及重要粮食和经济作物中鉴定出许多参与木聚糖生物合成的基因。该文综述了相关研究进展, 并探讨了木聚糖生物合成基因在生物质能及相关领域的潜在应用价值。
View image in article
图1
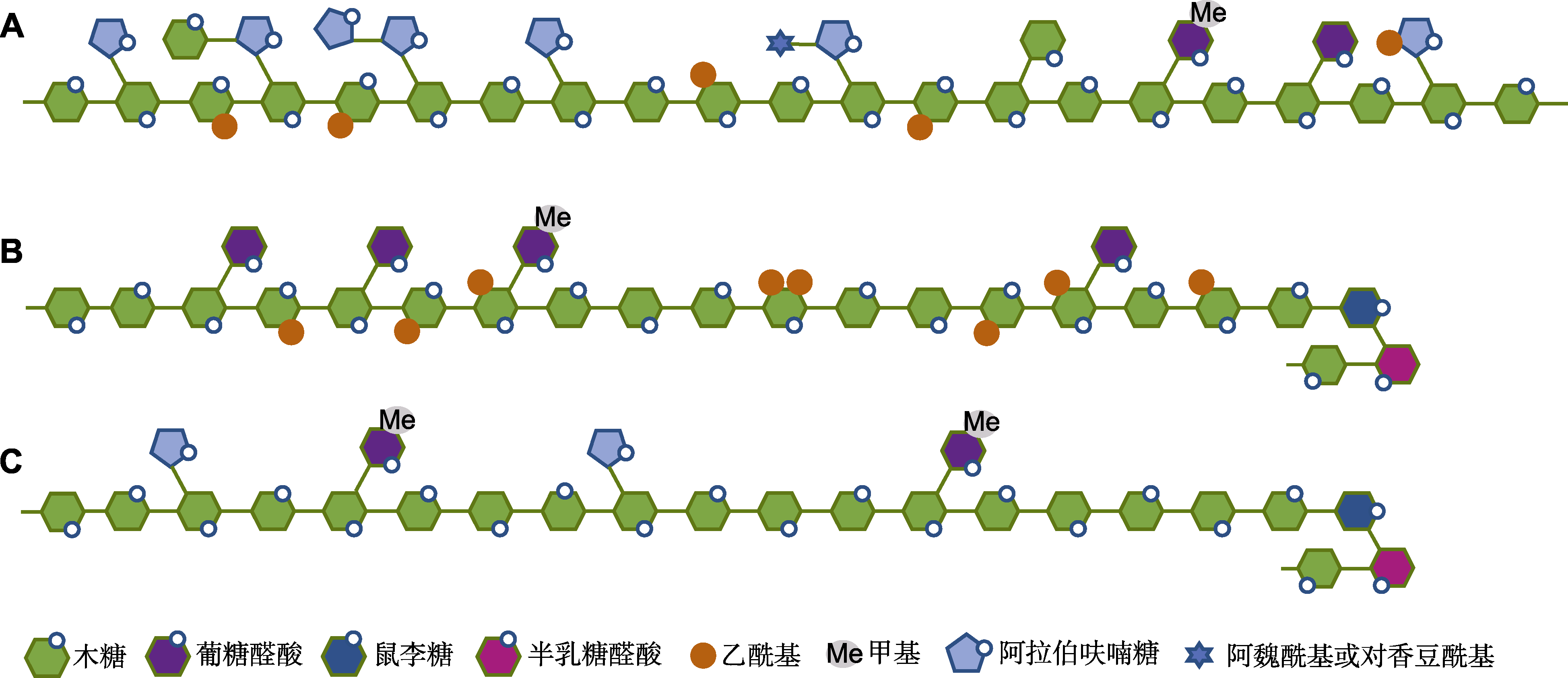
木聚糖结构示意图(参考Zhong et al., 2019; Zhang et al., 2021)
(A) 单子叶禾本科植物中的葡糖醛酸阿拉伯木聚糖; (B) 双子叶植物及一些非禾本科单子叶植物中的葡糖醛酸木聚糖; (C) 裸子植物中的葡糖醛酸阿拉伯木聚糖
正文中引用本图/表的段落
木聚糖的主链绝大多数是由木糖(xylose, Xyl)通过β-1,4糖苷键连接成的糖链(Pe?a et al., 2007), 在某些藻类中发现了β-1,4或β-1,4-1,3混合连接的主链(Deniaud et al., 2003)。根据主链上取代基的不同, 可将木聚糖分为3种, 即葡糖醛酸木聚糖(glucuronoxylan, GX)、阿拉伯木聚糖(arabinoxylan, AX)和葡糖醛酸阿拉伯木聚糖(glucuronoarabinoxylan, GAX) (图1)。葡糖醛酸木聚糖主要存在于双子叶植物以及一些非禾本科单子叶植物的次生壁中(Teleman et al., 2000; Pe?a et al., 2016)。阿拉伯木聚糖主要存在于单子叶禾本科植物的谷粒中(McCleary et al., 2015; Pellny et al., 2020)。葡糖醛酸阿拉伯木聚糖存在于双子叶植物的初生壁以及单子叶禾本科植物的营养组织和裸子植物(除买麻藤门外)的细胞壁中(Verbruggen et al., 1998; Busse-Wicher et al., 2016)。
ND: 尚未确定. ... The biosynthesis of L-arabinose in plants: molecular cloning and characterization of a Golgi-localized UDP-D-xylose 4-epimerase encoded by the MUR4 gene of Arabidopsis 1 2003 ... 高尔基体中的UDP-Xyl通过高尔基体定位的UDP-Xyl差向异构酶(UDP-Xyl epimerase, UXE)合成UDP-arabinopyranose (Arap) (Burget et al., 2003).UDP-Arap可能通过转运蛋白进入细胞质, 在定位于细胞质的UDP-Ara变位酶(UDP-Ara mutase, UAM)催化下合成UDP-Araf, 随后通过UDP-Araf转运蛋白(UDP-Araf transporter, UAfT)运送到高尔基体, 用于木聚糖上阿拉伯糖基取代(Rautengarten et al., 2017).用于木聚糖乙酰基取代的底物是乙酰辅酶A.乙酰辅酶A来源广泛, 它可以在不同的细胞器(如微体和线粒体)中合成并转移到胞质, 胞质中的乙酰辅酶A可能通过RWA (reduced wall acetylation)蛋白转移至高尔基体(Manabe et al., 2013). ... Evolution of xylan substitution patterns in gymnosperms and angiosperms: implications for xylan interaction with cellulose 2 2016 ... 木聚糖的主链绝大多数是由木糖(xylose, Xyl)通过β-1,4糖苷键连接成的糖链(Pe?a et al., 2007), 在某些藻类中发现了β-1,4或β-1,4-1,3混合连接的主链(Deniaud et al., 2003).根据主链上取代基的不同, 可将木聚糖分为3种, 即葡糖醛酸木聚糖(glucuronoxylan, GX)、阿拉伯木聚糖(arabinoxylan, AX)和葡糖醛酸阿拉伯木聚糖(glucuronoarabinoxylan, GAX) (图1).葡糖醛酸木聚糖主要存在于双子叶植物以及一些非禾本科单子叶植物的次生壁中(Teleman et al., 2000; Pe?a et al., 2016).阿拉伯木聚糖主要存在于单子叶禾本科植物的谷粒中(McCleary et al., 2015; Pellny et al., 2020).葡糖醛酸阿拉伯木聚糖存在于双子叶植物的初生壁以及单子叶禾本科植物的营养组织和裸子植物(除买麻藤门外)的细胞壁中(Verbruggen et al., 1998; Busse-Wicher et al., 2016). ...
ND: 尚未确定. ... Structural studies of the mix-linked β(1→3)/β(1→4)-D-xylans from the cell wall of Palmaria palmata (Rhodophyta 1 2003 ... 木聚糖的主链绝大多数是由木糖(xylose, Xyl)通过β-1,4糖苷键连接成的糖链(Pe?a et al., 2007), 在某些藻类中发现了β-1,4或β-1,4-1,3混合连接的主链(Deniaud et al., 2003).根据主链上取代基的不同, 可将木聚糖分为3种, 即葡糖醛酸木聚糖(glucuronoxylan, GX)、阿拉伯木聚糖(arabinoxylan, AX)和葡糖醛酸阿拉伯木聚糖(glucuronoarabinoxylan, GAX) (图1).葡糖醛酸木聚糖主要存在于双子叶植物以及一些非禾本科单子叶植物的次生壁中(Teleman et al., 2000; Pe?a et al., 2016).阿拉伯木聚糖主要存在于单子叶禾本科植物的谷粒中(McCleary et al., 2015; Pellny et al., 2020).葡糖醛酸阿拉伯木聚糖存在于双子叶植物的初生壁以及单子叶禾本科植物的营养组织和裸子植物(除买麻藤门外)的细胞壁中(Verbruggen et al., 1998; Busse-Wicher et al., 2016). ... Hydroxycinnamic acid-modified xylan side chains and their cross-linking products in rice cell walls are reduced in the Xylosyl arabinosyl substitution of xylan 1mutant 2 2022 ... 一些GT61成员具有木糖基转移酶活性.例如, 禾本科中XYXT (xylan xylosyltransferase)蛋白OsXYXT1、ZmXYXT1、SbXYXT1、BdXYXT1和PvXYXT1/ 2均可将木糖残基添加到木聚糖主链上, 形成木糖侧链(Zhong et al., 2018b, 2021).水稻OsXAX1 (xylosyl arabinosyl substitution of xylan 1)介导木糖向阿拉伯糖基侧链的添加.有趣的是, 水稻xax1突变体中羟基肉桂酸的含量也减少(Chiniquy et al., 2012).通过详细比较xax1突变体中含有羟基肉桂酸组分的木聚糖侧链的含量, 表明OsXAX1的功能是将被羟基肉桂酸修饰的阿拉伯糖基添加到木聚糖主链, 而不是将木糖添加到阿拉伯糖基侧链(Feijao et al., 2022). ...
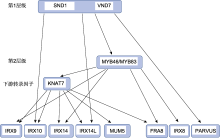
ND: 尚未确定. ... NAC-MYB-based transcriptional regulation of secondary cell wall biosynthesis in land plants 1 2015 ... 植物次生壁的生物合成由NAC及MYB等转录因子组成的层级网络所调控.第1层级是NAC类转录因子, 如NST1、SND1/NST3、VND6和VND7; 第2层级是MYB类转录因子MYB46/MYB83, 它们可以直接或间接地调控次生壁的合成, 是主开关转录因子, 在其下游还有一些转录因子, 特异性地调控某些基因的表达(Nakano et al., 2015; 张雨等, 2020).木聚糖作为次生壁半纤维素的主要成分, 其合成受该层级网络中的转录因子调控.已有证据表明, 位于第1层级的SND1和VND7、第1层级的直接靶标MYB46/MYB83以及MYB46的直接靶标KNAT7可以调控拟南芥木聚糖的合成(图3). ... Designing xylan for improved sustainable biofuel production 1 2019 ... 木聚糖是细胞壁中主要的半纤维素, 可以通过氢键与纤维素结合, 通过氢键和共价键与木质素和果胶结合, 参与细胞壁的形成, 决定细胞壁的物理性质(Tan et al., 2013; Broxterman and Schols, 2018; Kang et al., 2019; Terrett and Dupree, 2019; Gao et al., 2020a).木聚糖与人类的日常生活息息相关.例如, 黑麦(Secale cereale)和小麦(Triticum aestivum)中的阿拉伯木聚糖影响面包的加工工艺、品质和营养价值等(宋晓庆和董海洲, 2004); 玉米纤维麸皮阿拉伯木聚糖可作为食品添加剂、加厚剂、稳定剂、成膜剂及乳化剂(刘妍等, 2015).作为被子植物次生壁中含量仅次于纤维素的多糖, 木聚糖对植物的生长发育至关重要.许多研究表明, 严重缺乏木聚糖的拟南芥(Arabidopsis thaliana)突变体生长受阻, 细胞壁中木糖含量降低, 表现不规则木质部(irregular xylem, IRX)的表型, 而且植株异常矮小, 其原因可能是木聚糖的减少影响植物次生壁中纤维素微纤维的正确捆绑和排列(Brown et al., 2007; Crowe et al., 2021).在生物能源领域, 木聚糖与纤维素微纤丝的交联互作可降低纤维素酶对纤维素的可及性, 阻碍纤维素的降解(Smith et al., 2017).近年来, 越来越多的研究表明, 通过操纵木聚糖生物合成相关基因来改变细胞壁中木聚糖的含量或结构, 可以改变细胞壁的性质, 从而对植物的生长发育或生物质的加工特性产生影响(Smith et al., 2017; Oliveira et al., 2019; Bryant et al., 2020; Qaseem and Wu, 2020).因此, 全面了解木聚糖生物合成机制, 有助于利用分子生物学技术改良作物品质, 满足现代社会的发展需求. ... In muro deacetylation of xylan affects lignin properties and improves saccharification of aspen wood 1 2017 ... 将从黑曲霉(Aspergillus niger)中分离出的木聚糖乙酰酯酶基因AnAXE1转入杂种白杨(P. tremula × tremuloides), 可适度降低杂种白杨细胞壁中木聚糖的乙酰化程度, 使糖化作用得到增强而植物生长不受影响(Pawar et al., 2017).因此, 若以木材生物质作为第2代生物能源目标, 可以适当降低乙酰化程度.上述单子叶禾本科植物和木本植物木聚糖合成相关基因及应用详见表1. ... Acetylation of woody lignocellulose: significance and regulation 1 2013 ... 硬木木聚糖主链的C-2和/或C-3位置有70%会发生乙酰化, 而软木木聚糖主链通常缺乏乙酰化(Pawar et al., 2013).已证实毛果杨中有12个基因(PtrXOAT1- 12)参与调控木聚糖乙酰化的乙酰基转移酶(Zhong et al., 2018c).木聚糖乙酰化可以提高植物耐热性、机械强度和疏水性(de Carvalho et al., 2020).因此, 适度提高木本植物木聚糖的乙酰化程度可使木材更适用于建筑及家具制造等行业. ... Cell walls of developing wheat starchy endosperm: comparison of composition and RNA-seq transcriptome 1 2012 ... 小麦胚乳细胞壁的主要成分是阿拉伯木聚糖, 约占细胞壁总多糖的70%.小麦籽粒水提物(water-extractable, WE-AX)是饮食结构中可溶性膳食纤维的来源, 也是酿酒工业的重要原料.WE-AX黏度是影响各种加工过程的关键性状, 其很大程度取决于胚乳AX的性质.在小麦胚乳中表达量最高的IRX14、IRX9和IRX10的同源基因分别为TaGT43_1、TaGT43_2和TaGT47_2, 它们是胚乳中最有可能参与木聚糖主链合成的基因(Pellny et al., 2012).通过分析从小麦黄化幼苗中纯化的具有木聚糖合酶活性的蛋白, 发现TaGT43_1 (TaGT43-4)是该木聚糖合酶复合物的关键组分(Jiang et al., 2016).下调TaGT43_1、TaGT43_ 2或TaGT47_2对小麦胚乳AX产生显著影响, 使AX的数量减少且链长变短, WE-AX黏度降低, 更易提取(Freeman et al., 2016; Wilkinson et al., 2021).Pellny等(2020)证实TaGT43_2在小麦基因组中对应的3个同源基因TaGT43_2A、TaGT43_2B和TaGT43_ 2D (均命名为TaIRX9b)功能冗余地参与小麦胚乳AX主链合成.在TaIRX9b功能缺失三突变体小麦品系的胚乳中, AX含量降至对照的65%, 分子量减小且链长变短, WE-AX黏度降低50%-80%, 更易提取, 但包括胚乳在内的所有细胞的大小和形状并未发生改变, 这是酿酒工业所追求的优良性状(Pellny et al., 2020). ... Loss of TaIRX9b gene function in wheat decreases chain length and amount of arabinoxylan in grain but increases cross-linking 4 2020 ... 木聚糖的主链绝大多数是由木糖(xylose, Xyl)通过β-1,4糖苷键连接成的糖链(Pe?a et al., 2007), 在某些藻类中发现了β-1,4或β-1,4-1,3混合连接的主链(Deniaud et al., 2003).根据主链上取代基的不同, 可将木聚糖分为3种, 即葡糖醛酸木聚糖(glucuronoxylan, GX)、阿拉伯木聚糖(arabinoxylan, AX)和葡糖醛酸阿拉伯木聚糖(glucuronoarabinoxylan, GAX) (图1).葡糖醛酸木聚糖主要存在于双子叶植物以及一些非禾本科单子叶植物的次生壁中(Teleman et al., 2000; Pe?a et al., 2016).阿拉伯木聚糖主要存在于单子叶禾本科植物的谷粒中(McCleary et al., 2015; Pellny et al., 2020).葡糖醛酸阿拉伯木聚糖存在于双子叶植物的初生壁以及单子叶禾本科植物的营养组织和裸子植物(除买麻藤门外)的细胞壁中(Verbruggen et al., 1998; Busse-Wicher et al., 2016). ...
ND: 尚未确定. ... Structural diversity of xylans in the cell walls of monocots 3 2016 ... 木聚糖的主链绝大多数是由木糖(xylose, Xyl)通过β-1,4糖苷键连接成的糖链(Pe?a et al., 2007), 在某些藻类中发现了β-1,4或β-1,4-1,3混合连接的主链(Deniaud et al., 2003).根据主链上取代基的不同, 可将木聚糖分为3种, 即葡糖醛酸木聚糖(glucuronoxylan, GX)、阿拉伯木聚糖(arabinoxylan, AX)和葡糖醛酸阿拉伯木聚糖(glucuronoarabinoxylan, GAX) (图1).葡糖醛酸木聚糖主要存在于双子叶植物以及一些非禾本科单子叶植物的次生壁中(Teleman et al., 2000; Pe?a et al., 2016).阿拉伯木聚糖主要存在于单子叶禾本科植物的谷粒中(McCleary et al., 2015; Pellny et al., 2020).葡糖醛酸阿拉伯木聚糖存在于双子叶植物的初生壁以及单子叶禾本科植物的营养组织和裸子植物(除买麻藤门外)的细胞壁中(Verbruggen et al., 1998; Busse-Wicher et al., 2016). ...
ND: 尚未确定. ... An Arabidopsis cell wall proteoglycan consists of pectin and arabinoxylan covalently linked to an arabinogalactan protein 1 2013 ... 木聚糖是细胞壁中主要的半纤维素, 可以通过氢键与纤维素结合, 通过氢键和共价键与木质素和果胶结合, 参与细胞壁的形成, 决定细胞壁的物理性质(Tan et al., 2013; Broxterman and Schols, 2018; Kang et al., 2019; Terrett and Dupree, 2019; Gao et al., 2020a).木聚糖与人类的日常生活息息相关.例如, 黑麦(Secale cereale)和小麦(Triticum aestivum)中的阿拉伯木聚糖影响面包的加工工艺、品质和营养价值等(宋晓庆和董海洲, 2004); 玉米纤维麸皮阿拉伯木聚糖可作为食品添加剂、加厚剂、稳定剂、成膜剂及乳化剂(刘妍等, 2015).作为被子植物次生壁中含量仅次于纤维素的多糖, 木聚糖对植物的生长发育至关重要.许多研究表明, 严重缺乏木聚糖的拟南芥(Arabidopsis thaliana)突变体生长受阻, 细胞壁中木糖含量降低, 表现不规则木质部(irregular xylem, IRX)的表型, 而且植株异常矮小, 其原因可能是木聚糖的减少影响植物次生壁中纤维素微纤维的正确捆绑和排列(Brown et al., 2007; Crowe et al., 2021).在生物能源领域, 木聚糖与纤维素微纤丝的交联互作可降低纤维素酶对纤维素的可及性, 阻碍纤维素的降解(Smith et al., 2017).近年来, 越来越多的研究表明, 通过操纵木聚糖生物合成相关基因来改变细胞壁中木聚糖的含量或结构, 可以改变细胞壁的性质, 从而对植物的生长发育或生物质的加工特性产生影响(Smith et al., 2017; Oliveira et al., 2019; Bryant et al., 2020; Qaseem and Wu, 2020).因此, 全面了解木聚糖生物合成机制, 有助于利用分子生物学技术改良作物品质, 满足现代社会的发展需求. ... Characterization of acetylated 4-O-methylglucuronoxylan isolated from aspen employing 1H and 13C NMR spectroscopy 2 2000 ... 木聚糖的主链绝大多数是由木糖(xylose, Xyl)通过β-1,4糖苷键连接成的糖链(Pe?a et al., 2007), 在某些藻类中发现了β-1,4或β-1,4-1,3混合连接的主链(Deniaud et al., 2003).根据主链上取代基的不同, 可将木聚糖分为3种, 即葡糖醛酸木聚糖(glucuronoxylan, GX)、阿拉伯木聚糖(arabinoxylan, AX)和葡糖醛酸阿拉伯木聚糖(glucuronoarabinoxylan, GAX) (图1).葡糖醛酸木聚糖主要存在于双子叶植物以及一些非禾本科单子叶植物的次生壁中(Teleman et al., 2000; Pe?a et al., 2016).阿拉伯木聚糖主要存在于单子叶禾本科植物的谷粒中(McCleary et al., 2015; Pellny et al., 2020).葡糖醛酸阿拉伯木聚糖存在于双子叶植物的初生壁以及单子叶禾本科植物的营养组织和裸子植物(除买麻藤门外)的细胞壁中(Verbruggen et al., 1998; Busse-Wicher et al., 2016). ...
ND: 尚未确定. ... Xyloglucan O-acetyltransferases from Arabidopsis thaliana and Populus trichocarpa catalyze acetylation of fucosylated galactose residues on xyloglucan side chains 1 2018d ... 乙酰化是一种广泛存在于细胞壁聚合物中的修饰方式, 木葡聚糖、木聚糖及果胶等多聚物上均存在乙酰化修饰, 且修饰位点具有多样性(Gille and Pauly, 2012).因此需要众多乙酰化酶和去乙酰化酶参与调控.目前发现3类参与木聚糖乙酰化的基因, 分别是RWA、TBL (trichome birefringence-like)和AXY9 (altered xyloglucan 9).拟南芥中有4个RWA类基因(RWA1-4), 它们可能编码转运蛋白, 将乙酰辅酶A从细胞质转运至高尔基体, 但是其活性有待检测(Lee et al., 2011a; Manabe et al., 2013).TBL类蛋白直接催化木聚糖乙酰化, 特异地向主链木糖残基的2-O和(或) 3-O位点添加乙酰基.拟南芥中有9个参与木聚糖乙酰化的TBL蛋白, 即TBL3和TBL28-35, 它们均含有TBL结构域和DUF231结构域(Yuan et al., 2016a, 2016b; Zhong et al., 2017a).TBL29 (又称ESK1或XOAT1)是第1个被报道参与木聚糖乙酰化的基因.体外实验证实TBL29是乙酰基转移酶(Xiong et al., 2013; Yuan et al., 2013; Urbanowicz et al., 2014).Lunin等(2020)证明TBL29只能催化木糖残基的2-O位点发生乙酰化, 2-O位点的乙酰基可以迁移到3-O位点, 从而在2-O和3-O位点都产生单乙酰化.不同于RWA和TBL, AXY9是一个特殊的乙酰化相关酶, 它在多糖乙酰化中具有广泛作用.axy9突变体中木聚糖和木葡聚糖的乙酰基含量均有所下降(Schultink et al., 2015).目前尚不清楚AXY9的作用机理.生化实验显示, 重组的AXY9无乙酰基转移酶活性, 推测它可能是将乙酰基从乙酰辅酶A转移至TBL蛋白的媒介(Zhong et al., 2018d). ... Secondary cell wall biosynthesis 3 2019 ... 木聚糖的主链绝大多数是由木糖(xylose, Xyl)通过β-1,4糖苷键连接成的糖链(Pe?a et al., 2007), 在某些藻类中发现了β-1,4或β-1,4-1,3混合连接的主链(Deniaud et al., 2003).根据主链上取代基的不同, 可将木聚糖分为3种, 即葡糖醛酸木聚糖(glucuronoxylan, GX)、阿拉伯木聚糖(arabinoxylan, AX)和葡糖醛酸阿拉伯木聚糖(glucuronoarabinoxylan, GAX) (图1).葡糖醛酸木聚糖主要存在于双子叶植物以及一些非禾本科单子叶植物的次生壁中(Teleman et al., 2000; Pe?a et al., 2016).阿拉伯木聚糖主要存在于单子叶禾本科植物的谷粒中(McCleary et al., 2015; Pellny et al., 2020).葡糖醛酸阿拉伯木聚糖存在于双子叶植物的初生壁以及单子叶禾本科植物的营养组织和裸子植物(除买麻藤门外)的细胞壁中(Verbruggen et al., 1998; Busse-Wicher et al., 2016).
本文的其它图/表
|

 首页
首页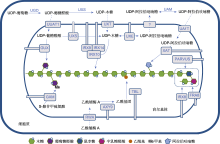
{kind=link}