|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
表观遗传调控植物分枝/分蘖研究进展
植物学报
2022, 57 (4):
532-548.
DOI: 10.11983/CBB22027
分蘖是禾本科植物特有的分枝类型, 是影响作物产量的关键因素之一。分枝/分蘖数由叶腋处侧生分生组织的数量和侧芽的活性共同决定。表观遗传修饰调控植物生长发育的各个方面, 但是如何调控植物的分枝/分蘖数还未见系统报道。该综述归纳了表观遗传调控侧生分生组织的形成和侧芽向外生长两个方面, 并展望了表观遗传在调控植物分枝/分蘖中的研究方向, 以期为通过表观遗传修饰改良作物品种的育种途径提供理论指导。
表2
表观遗传调控植物分枝/分蘖基因
正文中引用本图/表的段落
植物的表型具有很强的可塑性。株型是许多作物产量的决定因素之一。株型由植株高度和分枝数量等因素决定。分蘖是水稻等禾本科植物特有的分枝。分枝/分蘖数目主要由2个因素决定, 即侧生分生组织(axillary meristem, AM)的数量和侧芽的活性, 二者又受外部环境和体内激素的共同调控。
表观遗传参与生命过程的各个环节, 但其调控植物分枝/分蘖的报道相对较少。本文综述了近年来植物侧芽形成和伸长的最新研究进展, 以及表观遗传在这2个过程中的调控作用, 并对今后其在调控植物分枝/分蘖中的研究重点进行展望。
拟南芥中存在3个功能冗余的RAX (REGULATOR OF AXILLARY MERISTEMS)基因, 编码R2R3 MYB转录因子, 共同调控AM的形成。RAX2和RAX3主要在叶腋的中心区表达。rax1突变体有最强的AM发生缺陷表型, RAX2和RAX3突变能加重rax1的表型, rax1/rax2/rax3三突变体的莲座叶和茎生叶都很少产生侧芽(Mu?ller et al., 2006)。番茄RAX的同源基因Bl (Blind)也影响番茄侧芽的数量(Schmitz et al., 2002); 水稻RAX的同源基因RGN1 (REGULATOR OF GRAIN NUMBER 1)调控水稻穗型和每穗粒数(Li et al., 2022)。
拟南芥中有3个CUC (CUP-SHAPED COTYLEDON)基因, 编码NAC转录因子, 三者通过功能冗余的方式影响拟南芥AM的形成(Aida et al., 1997; Hibara et al., 2006)。cuc1/cuc2双突变体偶尔会出现杯状子叶(Aida et al., 1999), cuc3突变体表现出明显的莲座叶处AM缺陷, CUC1和CUC2突变能够加重cuc3的表型, 三突变体的莲座叶和茎生叶处均无侧芽(Hibara et al., 2006; Raman et al., 2008)。由于表达部位不同, 只有CUC1以及CUC2的转录本受miR164调控(Raman et al., 2008)。CUC2的启动子区有大量的转录因子识别位点, 故被认为是侧芽形成的重要调控枢纽(Tian et al., 2014)。CUC调控分生组织标记基因STM和LAS的表达。在cuc1/cuc2双突变体心形胚胎发育时期检测不到STM的表达。STM蛋白也可结合CUC1的启动子并激活其表达, 形成正向反馈环(Raman et al., 2008; Tian et al., 2014; Scofield et al., 2018)。Tian等(2014)研究发现, 在cuc1/cuc2/ cuc3三突变体的叶腋处检测不到LAS的表达, CUC2可直接结合LAS的启动子区调控其表达。
表观遗传修饰允许相同的基因组在不同的细胞类型中表现出不同的表达模式。表观遗传几乎参与植物所有的发育过程, 包括AM的形成。干细胞标记基因WUS和分生组织标记基因STM的转录调控均需要宽松的表观遗传环境, 以确保其精确地时空表达。多梳蛋白复合体PRC (polycomb repressive complex)调控靶基因H3K27me3组蛋白修饰并抑制基因的表达(Mozgova and Hennig, 2015; Förderer et al., 2016)。在成熟的器官(如叶片)中STM和WUS均有较高水平的H3K27me3标记, 而在含有STM表达的叶腋组织中, 此2个基因的H3K27me3水平降低, 而与染色质激活状态相关的标记H3Ac水平升高(Shi et al., 2016; Wang et al., 2017)。在PRC相关突变体中, WUS以及STM的表达量均被上调(Schubert et al., 2006; Liu et al., 2011; Shi et al., 2016; Wang et al., 2017)。此外, 用组蛋白脱乙酰化抑制剂trichostatin A处理成熟叶片可导致WUS异位表达(Wang et al., 2017)。表观遗传对WUS的调控是动态的。在幼叶的叶腋处WUS不表达, 此时H3K27me3标记比较丰富, 几乎检测不到H3Ac和H4Ac修饰。在WUS被激活前, 抑制标记H3K27me3减少, 而激活标记H3/4Ac增加(Wang et al., 2017)。
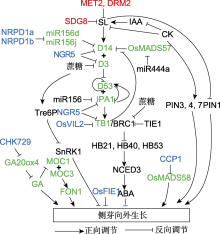
TCP转录因子包含bHLH结构域, 该结构域可结合DNA及介导蛋白互作。研究发现BRC1可结合并激活3个HD-ZIP I转录因子(HB21、HB40和HB53), 这3个转录因子基因可促进脱落酸(ABA)生物合成基因NCED3 (9-CIS-EPOXICAROTENOID DIOXYGENASE 3)的表达, 导致ABA水平升高(图2), 而ABA能够增强侧芽的休眠(González-Grandío et al., 2017)。这些HD-ZIP I转录因子基因可能是BRC1调控侧芽向外生长的靶基因。此外, Van Es等(2020)发现, BRC1对ABA信号具有更广泛的调控作用, 可直接调控参与ABA代谢、运输、感知、信号转导和响应的基因表达。
由AM形成的侧芽受顶端分生组织的调控而维持休眠状态, 该现象称为顶端优势。研究表明IAA、SL、糖及环境因素均能影响顶端优势(图2)。目前, 侧芽打破休眠向外生长主要有两种假说, 分别是营养假说和IAA运输渠化假说(Phillips, 1975; Balla et al., 2011, 2016)。
目前, 有关表观遗传调控植物侧芽生长的报道主要在以下几个方面(表2; 图2)。
H3K27me3标记由Polycomb蛋白家族介导。PRC1和PRC2是2个调控基因表达的主要复合体。PRC2具有甲基转移酶活性, 对靶基因进行H3K27me3标记; PRC1具有泛素连接酶活性, 对H2A上第119个赖氨酸进行单泛素化修饰(H2AK119ub) (Ringrose and Paro, 2004)。通常认为PRC2复合体结合到特定的DNA序列PRE (polycomb response elements)上, 并对附近的DNA进行H3K27me3标记, 随后招募PRC1复合体进行H2AK119ub标记, 通过染色质重塑维持基因上H3K27me3标记(Berger et al., 2007; Endoh et al., 2012)。
水稻染色质重塑因子OsVIL2调控靶基因的H3K27me3标记水平, 进而调节基因的表达。osvil2突变体分蘖变少, 解剖学观察发现基因突变后影响分蘖芽的生长, 而不是侧生分生组织的形成。通过分析分蘖相关基因的表达, 发现OsTB1在突变体中的表达量升高。OsVIL2能结合OsTB1的启动子区, osvil2突变体中由于H3K27me3修饰降低, 导致OsTB1的表达量升高, 从而影响水稻分蘖芽的生长(Yoon et al., 2019)。
本文的其它图/表
|
 首页
首页