|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
表观遗传调控植物分枝/分蘖研究进展
植物学报
2022, 57 (4):
532-548.
DOI: 10.11983/CBB22027
分蘖是禾本科植物特有的分枝类型, 是影响作物产量的关键因素之一。分枝/分蘖数由叶腋处侧生分生组织的数量和侧芽的活性共同决定。表观遗传修饰调控植物生长发育的各个方面, 但是如何调控植物的分枝/分蘖数还未见系统报道。该综述归纳了表观遗传调控侧生分生组织的形成和侧芽向外生长两个方面, 并展望了表观遗传在调控植物分枝/分蘖中的研究方向, 以期为通过表观遗传修饰改良作物品种的育种途径提供理论指导。
表1
调控侧生分生组织形成的转录因子
正文中引用本图/表的段落
通过遗传学方法鉴定出多个转录因子调控侧生分生组织的形成(表1; 图1)。
LAS (LATERAL SUPPRESSOR)是第1个被鉴定出来影响AM形成的调控因子, LAS基因在叶腋处特异表达。拟南芥(Arabidopsis thaliana) las突变体缺少AM, LAS在番茄(Solanum lycopersicum)中的同源基因LS (LATERAL SUPPRESSOR)和水稻中的同源基因MOC1 (MONOCULM 1)不但影响AM的发生, 也影响花发育。ls突变体的花瓣发育异常(Schumacher et al., 1999), moc1突变体的花序分枝发育异常(Li et al., 2003)。LS/LAS/MOC1基因编码GRAS家族转录因子, 但目前其下游基因仍不清楚(Schumacher et al., 1999; Li et al., 2003)。在细胞周期后期促进复合物的共激活子TAD1 (TILLERING AND DWARF 1)/TE (TILLER ENHANCER)识别MOC1, 以依赖细胞周期的方式对其泛素化, 并招募26S蛋白酶体进行降解(Lin et al., 2012; Xu et al., 2012)。MOC1可以与水稻DELLA蛋白SLR1 (SLENDER RICE 1)互作, 维持MOC1蛋白的稳定性。赤霉素(gibberellins, GAs)能诱发SLR1的降解, 导致水稻茎秆伸长, 也能促进MOC1降解, 降低水稻的分蘖数目(Liao et al., 2019)。
拟南芥中存在3个功能冗余的RAX (REGULATOR OF AXILLARY MERISTEMS)基因, 编码R2R3 MYB转录因子, 共同调控AM的形成。RAX2和RAX3主要在叶腋的中心区表达。rax1突变体有最强的AM发生缺陷表型, RAX2和RAX3突变能加重rax1的表型, rax1/rax2/rax3三突变体的莲座叶和茎生叶都很少产生侧芽(Mu?ller et al., 2006)。番茄RAX的同源基因Bl (Blind)也影响番茄侧芽的数量(Schmitz et al., 2002); 水稻RAX的同源基因RGN1 (REGULATOR OF GRAIN NUMBER 1)调控水稻穗型和每穗粒数(Li et al., 2022)。
拟南芥中有3个CUC (CUP-SHAPED COTYLEDON)基因, 编码NAC转录因子, 三者通过功能冗余的方式影响拟南芥AM的形成(Aida et al., 1997; Hibara et al., 2006)。cuc1/cuc2双突变体偶尔会出现杯状子叶(Aida et al., 1999), cuc3突变体表现出明显的莲座叶处AM缺陷, CUC1和CUC2突变能够加重cuc3的表型, 三突变体的莲座叶和茎生叶处均无侧芽(Hibara et al., 2006; Raman et al., 2008)。由于表达部位不同, 只有CUC1以及CUC2的转录本受miR164调控(Raman et al., 2008)。CUC2的启动子区有大量的转录因子识别位点, 故被认为是侧芽形成的重要调控枢纽(Tian et al., 2014)。CUC调控分生组织标记基因STM和LAS的表达。在cuc1/cuc2双突变体心形胚胎发育时期检测不到STM的表达。STM蛋白也可结合CUC1的启动子并激活其表达, 形成正向反馈环(Raman et al., 2008; Tian et al., 2014; Scofield et al., 2018)。Tian等(2014)研究发现, 在cuc1/cuc2/ cuc3三突变体的叶腋处检测不到LAS的表达, CUC2可直接结合LAS的启动子区调控其表达。
EXB1 (EXCESSIVE BRANCHES 1)编码WRKY71转录因子。过表达EXB1的拟南芥会产生较多的分枝。EXB1蛋白融合EAR结构域抑制下游基因的表达, 导致植物分枝减少, 而exb1突变体不具备分枝表型, 可能是由于功能冗余所致。EXB1可调控RAX1的表达并激活AM (Yang et al., 2012; Guo et al., 2015)。
玉米TB1 (TEOSINTE BRANCHED 1)是调控分枝的1个关键转录因子。在驯化过程中显性过表达TB1基因抑制玉米产生过多分枝(Doebley et al., 1995; Hubbard et al., 2002)。TB1在水稻中的同源基因OsTB1/FC1 (FINECULM 1)和拟南芥中的同源基因BRC1 (BRANCHED 1)均在休眠芽中特异表达。过表达TB1及其同源基因能够抑制分枝/分蘖, 相应的功能缺失突变体则表现出多分枝/多分蘖表型。因此, TB1/BRC1被认为是抑制侧芽向外生长所必需(Takeda et al., 2003; Minakuchi et al., 2010; González-Grandío et al., 2013)。但是, BRC1并不是抑制侧芽生长的唯一因子, 在brc1突变植株中, 并非所有芽均被释放出来长成分枝(Arite et al., 2007; Finlayson, 2007; Braun et al., 2012)。brc1和独脚金内酯(strigolactone, SL)生物合成或者信号转导突变体的双突变体表现出更多分枝的表型(Braun et al., 2012; Chevalier et al., 2014; Seale et al., 2017); 且植株对去除顶端、细胞分裂素和环境因素(如栽培密度)表现出一定的响应(Takeda et al., 2003; Ferguson and Beveridge, 2009; Braun et al., 2012; Dun et al., 2012; González-Grandío et al., 2013)。在缺少BRC1的情况下分枝也可被抑制, 在BRC1表达的情况下也可被激活(Seale et al., 2017)。
BRC1在蛋白水平被转录抑制因子TIE1调控。含有EAR结构域的TIE1可与BRC1互作, 表达BRC1融合TIE1碳末端蛋白可增加植物的分枝, 类似于过表达TIE1和brc1功能缺失突变体的多分枝表型, 说明BRC1蛋白活性的调控对植物分枝也很重要(Yang et al., 2018)。
SL促进BRC1表达但CK抑制其表达(Dun et al., 2012)。Mason等(2014)研究发现, 去掉或减少生长旺盛部位, 叶片中产生的糖可促进侧芽生长。外源施加蔗糖也可促使侧芽打破休眠, 并可抑制BRC1的表达。此外, 其它植物激素、营养供给和环境因子共同调控芽的向外生长, OsTB1/BRC1是芽能否被激活的重要调控靶点(Seale et al., 2017)。
MOC1和MOC3除了影响AM的形成, 还可调控侧芽的生长。FON1 (FLORAL ORGAN NUMBER 1)是CLV1 (CLAVATA 1)在水稻中的同源基因, 在侧生分生组织中高水平表达。FON1功能缺失突变体分蘖数减少, 主要是由于分蘖芽的伸长受到影响, 而分蘖芽的形成未受影响。MOC3能直接结合到FON1基因启动子区激活其表达, 而MOC1蛋白可与MOC3互作, 增强MOC3对FON1的激活作用, 进而促进分蘖芽的伸长(Shao et al., 2019)。
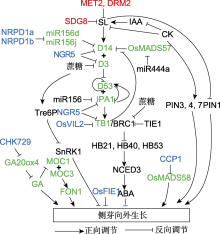
CK和SL被认为是调控侧芽生长的重要植物激素。CK促进芽的生长, 而SL起抑制作用。在顶端合成并在主茎中运输的IAA能够促进SL生物合成基因的表达(Domagalska and Leyser, 2011; Rameau et al., 2014)。与IAA不同, CK以及SL可进入侧芽(Teichmann and Muhr, 2015)。把CK和SL施加到主茎上可调控侧芽的生长(Dun et al., 2012), 且侧芽的向外生长与其中CK和SL信号基因的表达密切相关, 说明侧芽可能是SL和CK行使功能的场所(Mason et al., 2014; Dierck et al., 2016; Roman et al., 2016)。SL和CK被认为是通过控制侧芽中IAA的向外运输来调控侧芽生长(Crawford et al., 2010; Shinohara et al., 2013; Bennett et al., 2016; Waldie and Leyser, 2018)。事实上, SL能诱发IAA外排转运体PIN的胞吞(Shinohara et al., 2013; Rameau et al., 2014); 抑制IAA的生物合成(Ligerot et al., 2017)。max2的多分枝表型在PIN1、PIN3以及PIN7发生突变后得到部分恢复, 说明SL部分通过IAA的极性运输行使功能(Van Rongen et al., 2019)。但在SL缺陷豌豆突变体rms1中, IAA外排运输受到抑制后外源施加SL仍可以抑制侧芽的向外生长(Gomez-Roldan et al., 2008; Chabikwa et al., 2019), 说明SL抑制侧芽生长还有其它机制。在拟南芥CK信号相关突变体中, IAA外输转运体在质膜的极性分布降低, 说明CK促进IAA外输转运体的极性分布, 调控侧芽生长(Waldie and Leyser, 2018) (图2)。
水稻中D53可与SPL家族转录因子IPA1相互作用(Song et al., 2017)。IPA1转录因子能直接结合OsTB1的启动子区并激活其表达, 从而抑制水稻分蘖(Lu et al., 2013)。D53蛋白可与IPA1直接互作并抑制其转录活性, SL诱导D53蛋白降解后, IPA1激活下游基因的表达。此外, IPA1可直接结合D53的启动子反馈调节其表达(Song et al., 2017)。D53可以抑制自身的表达以及下游BRANCHED 1、TCP DOMAIN PROTEIN 1和PRODUCTION OF ANTHOCYANIN PIGMENT 1的表达, 分别调控突变体的分蘖、叶型和花青素含量(Wang et al., 2020) (图2)。
拟南芥中含有MBD (methyl-CpG-binding domain)结构域的AtMBD9突变后植株表现出多分枝和早花表型。AtMBD9能识别DNA甲基化的表观遗传信息, 整合DNA甲基化、染色质结构修饰和基因表达。开花负调节基因FLC在atmbd9突变体中的表达量降低, 过表达FLC的转基因植株恢复了atmbd9的早花表型但是未能恢复多分枝表型。通过分别与arx1和max4构建双突变体, 发现atmbd9调控的多分枝表型与生长素和SL信号调控通路不同, 其可能调控新的植物分枝途径(Peng et al., 2006)。
RNA聚合酶IV复合体的2个亚基OsNRPD1a和OsNRPD1b发生突变后, 可引起水稻矮化多分蘖表型。OsNRPD1a和OsNRPD1b为产生24 nt小干扰RNA (siRNA)所必需, 这些小干扰RNA在转座子(包括微型反向重复转座元件(miniature inverted repeat transposable element, MITEs )上引导DNA甲基化。在osnrpd1a/osnrpd1b双突变体中, 促进分蘖的OsMIR156d和OsMIR156j启动子区由于2个MITE位点的CHH甲基化水平发生改变, 导致在双突变体中其表达量升高; 抑制分蘖的D14基因由于其下游1个MITE位点的CHH甲基化水平变化, 导致其表达量相应降低(Xu et al., 2020)。
水稻染色质重塑因子OsVIL2调控靶基因的H3K27me3标记水平, 进而调节基因的表达。osvil2突变体分蘖变少, 解剖学观察发现基因突变后影响分蘖芽的生长, 而不是侧生分生组织的形成。通过分析分蘖相关基因的表达, 发现OsTB1在突变体中的表达量升高。OsVIL2能结合OsTB1的启动子区, osvil2突变体中由于H3K27me3修饰降低, 导致OsTB1的表达量升高, 从而影响水稻分蘖芽的生长(Yoon et al., 2019)。
MADS-box基因多在花器官发育方面起重要作用。CCP1 (CURVED CHIMERIC PALEA 1)基因突变后, 突变体表现出内稃发育异常和多分蘖表型。CCP1编码植物特异的EMF1 (EMBRYONIC FLOWER 1)类似蛋白, 其是PRC2核心蛋白成分。在ccp1突变体中OsMADS58的H3K27me3标记降低导致OsMADS58表达量升高, 进而引起内稃发育异常(Yan et al., 2015)。
FIE1 (FERTILIZATION-INDEPENDENT EN-DOSPERM 1)编码类似于Esc的蛋白, 其是PRC2另一个核心成员, 涉及靶基因的H3K27me3修饰, 抑制基因表达。Epi-df FIE1功能获得性突变体表现出株型变矮和分蘖数增多。FIE1可与PRC2复合体核心蛋白E(z)在水稻中的同源蛋白iEZ1互作, 可能通过形成PRC2复合体, 招募靶基因进行H3K27me3标记, 进而调节水稻的株高和分蘖(Zhang et al., 2012)。
植物侧芽的生长受SL、IAA和CK等多种植物激素与环境条件的影响。其中BRC1/OsTB1的表达被认为是调控侧芽生长的重要中心枢纽, 整合了多种植物激素和环境信号。但还存在一些不依赖于BRC1/OsTB1的其它调控机制影响侧芽生长, 这个过程是否受表观遗传调控, 有待进一步探索。
本文的其它图/表
|
 首页
首页