
|
|
||
|
基因枪介导的老芒麦遗传转化体系的建立
植物学报
2021, 56 (1):
62-70.
DOI: 10.11983/CBB20174
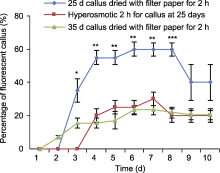
川草2号老芒麦(Elymus sibiricus)是青藏高原地区治理荒漠化和建设高产人工草地的主要栽培草种。用川草2号老芒麦5种外植体诱导愈伤组织, 经分化测试, 仅幼穗愈伤组织能分化再生。以当代培养25天和35天的结构致密坚硬的幼穗愈伤组织为受体, 分别进行农杆菌侵染和基因枪转化, 结果只有基因枪能转化成功。在基因枪转化过程中, 采用高渗培养和滤纸干燥2种方式预处理愈伤组织, 结果表明滤纸干燥处理比高渗处理转化效率高。当代诱导25天的幼穗愈伤组织, 滤纸干燥处理2小时转化效率最高, 达40%。该研究成功获得了基因枪转化的以川草2号老芒麦幼穗愈伤为受体的阳性愈伤组织。 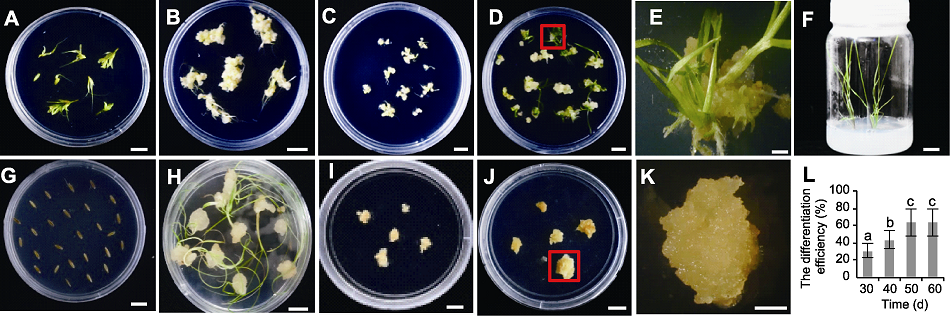 View image in article
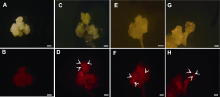
图4
川草2号老芒麦组培再生体系建立及幼穗愈伤分化效率随继代时间的变化
(A) 幼穗接种于愈伤诱导培养基; (B) 幼穗诱导35天后的愈伤; (C) 挑选图(B)中结构致密坚硬的愈伤置于继代培养基15天; (D) 图(C)愈伤置于分化培养基约35天; (E) 图(D)中红色框内愈伤分化显微图; (F) 图(D)中分化苗置于生根培养基约30天; (G) 种子接种于愈伤诱导培养基; (H) 种子诱导35天后的愈伤; (I) 挑选图(H)中结构致密坚硬的愈伤置于继代培养基15天; (J) 图(I)愈伤置于分化培养基约35天; (K) 图(J)中红色框内愈伤分化显微图; (L) 不同再生时间下的幼穗愈伤分化效率(误差线为标准误, 样本容量为30, 不同小写字母表示差异显著(P<0.05))。(D), (F), (G), (J) Bars=2 cm; (A)-(C), (E), (H), (I), (K) Bars=1 cm
正文中引用本图/表的段落
愈伤再生结果表明, 仅幼穗愈伤能分化出绿点和小苗(图4A-F)。幼穗愈伤在分化培养基继代30天后开始分化, 分化苗生根需25天, 从愈伤诱导到分化共需约115天。然而种子外植体诱导的愈伤无法获得再生苗(图4G-K), 根、茎、下胚轴均未成功建立再生体系(数据未列出)。在一定时间范围内, 幼穗愈伤分化率随再生时间推移而增加, 将幼穗愈伤置于分化培养基后约30天出现绿点, 分化率为30%; 50天后幼穗分化率不再增加, 达到峰值(约63%) (图4L)。
相比双子叶植物, 单子叶植物的遗传转化研究少且起步晚(杨静, 2019)。禾本科牧草愈伤再生困难(Vasil, 1994)是限制其遗传转化体系建立的主要因素。李达旭等(2006)用川草2号老芒麦种子(成熟胚)诱导出愈伤并建立遗传转化体系。本研究团队利用川草2号老芒麦约5万粒种子(成熟胚)诱导愈伤, 出现的愈伤均为海绵状且水渍化严重, 以该类型的愈伤进行分化实验, 均不能再生成完整植株或出现绿点(图4J)。推测实验结果不同是由于老芒麦种子的基因型不同所致。已有研究表明, 基因型是影响愈伤生长速率及分化再生的关键因素之一(Cheng et al., 2004)。禾本科牧草具自交不亲和及异源多倍体等遗传特性, 每粒种子均代表一个基因型(Wang and Ge, 2006; Li et al., 2006)。故来自每一粒种子的愈伤均可能属于不同基因型, 因此极有可能出现不同批次种子愈伤表现不同的情况, 这也是老芒麦遗传转化体系不稳定的主要原因之一。
本文的其它图/表
|
 首页
首页

{kind=link}