
植物学报 ›› 2020, Vol. 55 ›› Issue (6): 740-748.DOI: 10.11983/CBB20064 cstr: 32102.14.CBB20064
张春艳*( )
)
收稿日期:2020-04-20
接受日期:2020-08-26
出版日期:2020-11-01
发布日期:2020-11-11
通讯作者:
张春艳
作者简介:*E-mail: zhangchunyan@ibcas.ac.cn基金资助:
Chunyan Zhang*()
Received:2020-04-20
Accepted:2020-08-26
Online:2020-11-01
Published:2020-11-11
Contact:
Chunyan Zhang
摘要: P700氧化还原动力学技术可快速且无损地检测植物光系统I (PSI)的活性, 是光合研究领域中广泛使用的一种技术。该文系统归纳了P700氧化还原动力学的主要测量方法, 详细阐述其原理并探讨该技术的局限性, 旨在为深入研究光合作用机理提供技术支持。
张春艳. P700氧化还原动力学的测量方法及原理. 植物学报, 2020, 55(6): 740-748.
Chunyan Zhang. The Measurement Methods and Principles of P700 Redox Kinetics. Chinese Bulletin of Botany, 2020, 55(6): 740-748.
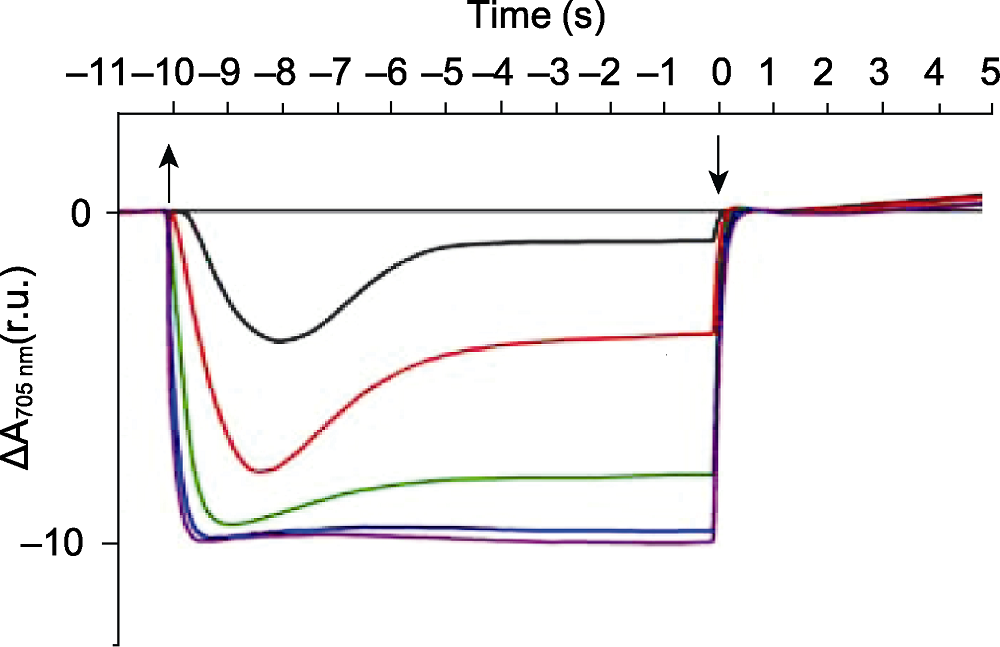
图1 活化光诱导的P700氧化还原动力学(改自Alric et al., 2010) 620 nm的红光光强为5 μmol·m-2·s-1 (黑色)、20 μmol·m-2·s-1 (红色)、50 μmol·m-2·s-1 (绿色)、200 μmol·m-2·s-1 (蓝色)、500 μmol·m-2·s-1 (紫色)。莱茵衣藻野生型加入10 μmol·L-1二氯苯基二甲脲(DCMU)和1 mmol·L-1羟胺(HA)。
Figure 1 The curves of P700 redox kinetics induced by actinic lights (modified from Alric et al., 2010) The red light intensity (620 nm) is approximately 5 μmol·m-2·s-1 (black), 20 μmol·m-2·s-1 (red), 50 μmol·m-2·s-1 (green), 200 μmol·m-2·s-1 (blue), 500 μmol·m-2·s-1 (purple). Wild type algae is incubated with 10 μmol·L-1 3-(3,4-dichloro- phenyl)-1,1-dimethylurea (DCMU) and 1 mmol·L-1 hydroxylamine (HA).
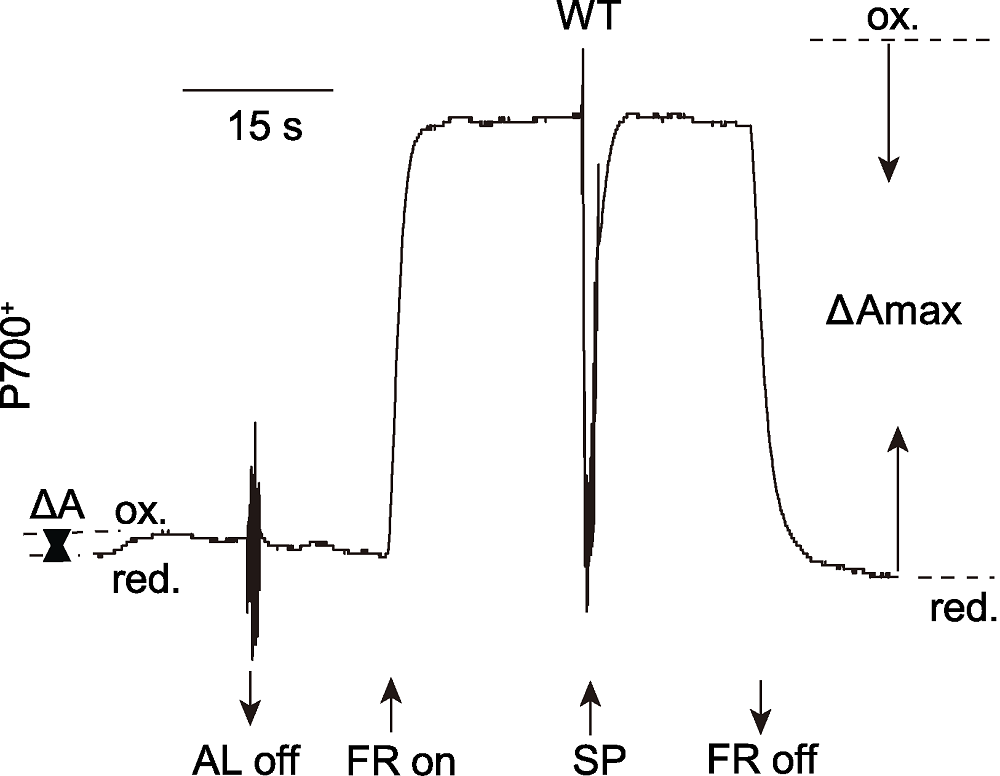
图2 饱和脉冲法计算PSI光化学量子产量Y(I) (改自Yang et al., 2017) AL: 活化光(80 μmol·m-2·s-1); FR: 远红光(720 nm, 20 μmol·m-2·s-1); SP: 饱和脉冲(3000 μmol·m-2·s-1); ΔA: 活化光诱导的部分氧化的P700; ΔAmax: 最大光氧化的P700; ox.: 氧化状态的P700; red.: 还原状态的P700
Figure 2 The saturation pulse method for determination of Y(I) (modified from Yang et al., 2017) AL: Actinic light (80 μmol·m-2·s-1); FR: Far-red light (720 nm, 20 μmol·m-2·s-1); SP: Saturation pulse (3000 μmol·m-2·s-1); ΔA: Partial photo-oxidizable P700 induced by AL; ΔAmax: Maximum photo-oxidizable P700; ox.: P700 oxidation; red.: P700 reduction
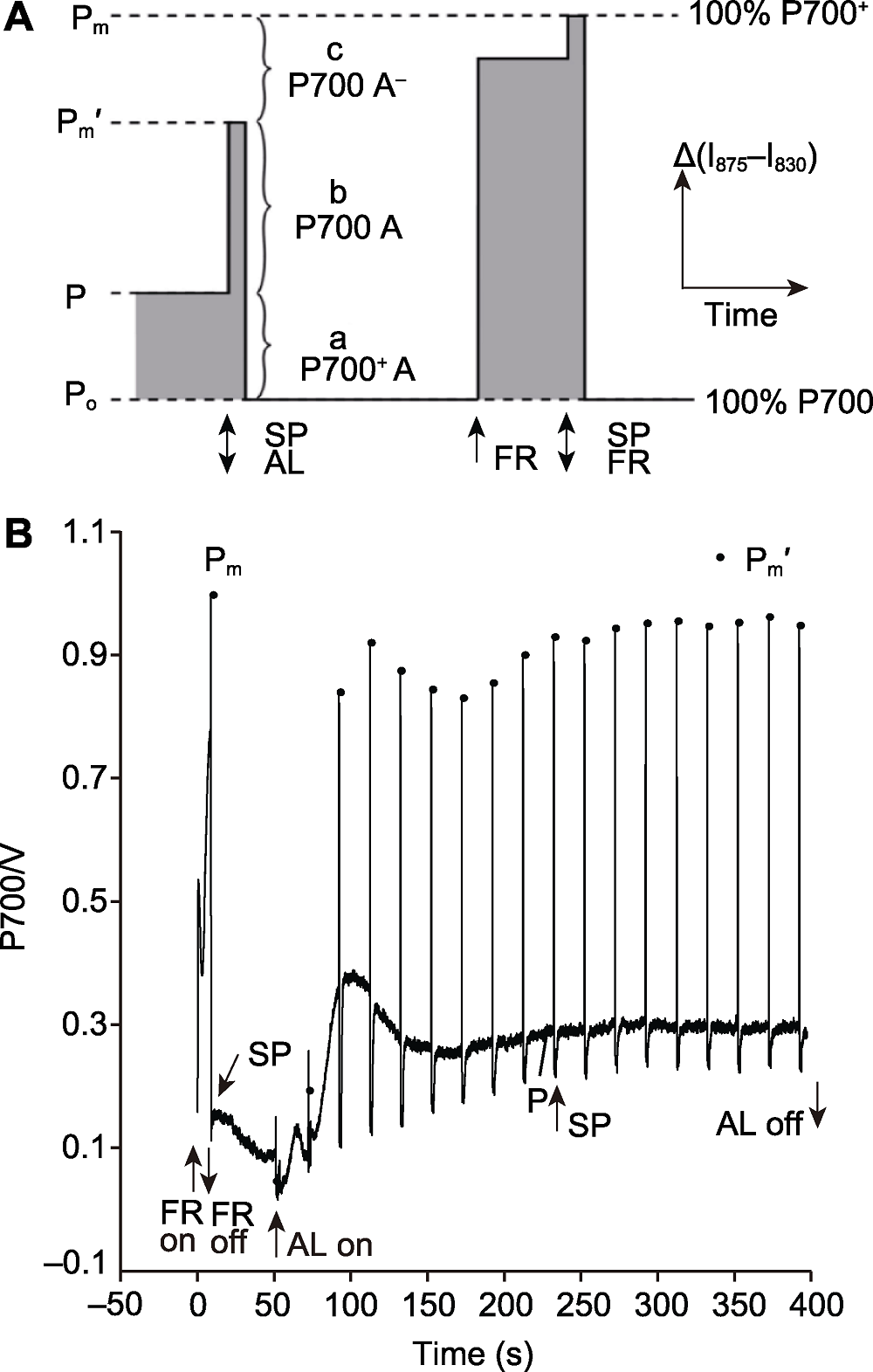
图3 利用饱和脉冲法计算PSI能量转化 (A)利用饱和脉冲法计算PSI能量转化的原理示意图(改自Klughammer and Schreiber, 2008); (B) 利用饱和脉冲法计算PSI能量转化的测量曲线。AL: 活化光; SP: 饱和脉冲(10000 μmol·m-2·s-1); FR: 远红光(720 nm); Po: 远红光和饱和脉冲关闭后, 全部PSI反应中心处于完全还原状态, 此时P700信号是0; P: 活化光照光过程中, 部分PSI反应中心被氧化的P700信号; Pm': 活化光关闭, 具有开放、有活性的PSI反应中心被饱和脉冲完全氧化的P700信号; Pm: 远红光关闭, 全部PSI反应中心被饱和脉冲完全氧化的P700信号; a P700+ A: 由于PSI电子供体侧限制导致的关闭的PSI反应中心; A: PSI的下游电子受体; b P700 A: 开放的PSI反应中心; c P700 A-: 由于PSI电子受体侧限制导致的关闭的PSI反应中心; a、b、c是PSI反应中心的3个部分, b是开放的、有活性的, 其光化学量子产量定义为1, 而a和c是关闭的、没有活性的, 其光化学量子产量定义为0。
Figure 3 The saturation pulse method for determination of efficiency of energy conversion in PSI (A) Principle of saturation pulse method for determination of efficiency of energy conversion in PSI (modified from Klughammer and Schreiber, 2008); (B) Curve of saturation pulse method for determination of efficiency of energy conversion in PSI. AL: Actinic light; SP: Saturation pulse (10000 μmol·m-2·s-1); FR: Far-red light (720 nm); Po: Complete reduction is induced after the cessation of SP and far-red light with the zero P700 signal; P: In the presence of AL, a part of the PSI centers are oxidized by the AL resulting in an intermediate P700 signal; Pm': A part of the PSI centers are oxidized completely by the SP with the maximal P700 signal after the AL is turned off; Pm: All the PSI centers are oxidized completely by the SP with the maximal P700 signal after FR pre-illumination; a P700+ A: Donor-side limited closed centers; A: The effective ensemble of PSI acceptors, the same as below; b P700 A: Open centers P700A; c P700 A-: Acceptor-side limited closed centers. a, b, c are the three parts of all PSI centers, b is open and active, and the photochemical quantum yield is unity, while a and c are closed and inactive, and the photochemical quantum yield is 0.
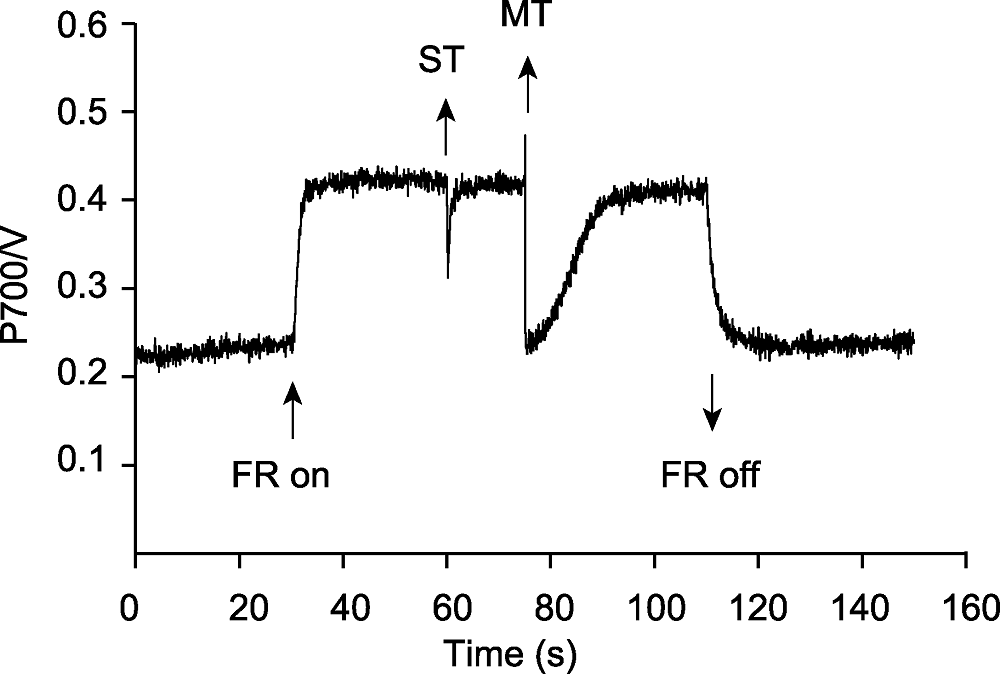
图4 光系统间的电子载体库 FR: 远红光(720 nm, 10 μmol·m-2·s-1); ST: 单周转饱和脉冲(200000 μmol·m-2·s-1); MT: 多周转饱和脉冲(20000 μmol·m-2·s-1)
Figure 4 The capacity of the intersystem electron carrier pool FR: Far-red light (720 nm, 10 μmol·m-2·s-1); ST: Single turnover flash (200000 μmol·m-2·s-1); MT: Multiple turnover flash (20000 μmol·m-2·s-1)
| [1] | 高坤山 (2014). 藻类固碳: 理论、进展与方法. 北京: 理科学出版社. pp. 426-427. |
| [2] | 沈同, 王镜岩, 赵邦悌, 李建武, 徐长法, 朱圣庚, 俞梅敏, 杨端, 杨福愉 (1991). 生物化学(第2版). 北京: 高等教育出版社. pp. 139. |
| [3] | 姚正菊, 米华玲, 叶济宇 (2002). 光诱导叶片P-700氧化还原的测量. 植物生理学通讯 38, 54-56. |
| [4] | 郁飞, 唐崇钦, 辛越勇, 彭德川, 许亦农, 李良璧, 匡廷云 (2001). 光系统I (PSI)的结构与功能研究进展. 植物学通报 18, 266-275. |
| [5] |
Alric J, Lavergne J, Rappaport F (2010). Redox and ATP control of photosynthetic cyclic electron flow in Chlamydomonas reinhardtii (I) aerobic conditions. Biochim Biophys Acta 1797, 44-51.
URL PMID |
| [6] |
Amunts A, Drory O, Nelson N (2007). The structure of a plant photosystem I supercomplex at 3.4 Å resolution. Nature 447, 58-63.
DOI URL PMID |
| [7] |
Amunts A, Nelson N (2009). Plant photosystem I design in the light of evolution. Structure 17, 637-650.
URL PMID |
| [8] | Andreeva A, Tikhonov AN (1983). Comparative study on the kinetics of electron transport and the slow chlorophyll fluorescence in bean leaves. Photobiochem Photobiophys 6, 261-266. |
| [9] | Asada K, Heber U, Schreiber U (1992). Pool size of electrons that can be donated to P700+ as determined in intact leaves: donation to P700+ from stromal components via the intersystem chain . Plant Cell Physiol 33, 927-932. |
| [10] |
Ballottari M, Alcocer MJP, D'Andrea C, Viola D, Ahn TK, Petrozza A, Polli D, Fleming GR, Cerullo G, Bassi R (2014). Regulation of photosystem I light harvesting by zeaxanthin. Proc Natl Acad Sci USA 111, E2431-E2438.
DOI URL PMID |
| [11] |
Bonente G, Pippa S, Castellano S, Bassi R, Ballottari M (2012). Acclimation of Chlamydomonas reinhardtii to different growth irradiances. J Biol Chem 287, 5833-5847.
URL PMID |
| [12] | Brettel K (1997). Electron transfer and arrangement of the redox cofactors in photosystem I. Biochim Biophys Acta 1318, 322-373. |
| [13] |
Brettel K, Leibl W (2001). Electron transfer in photosystem I. Biochim Biophys Acta 1507, 100-114.
DOI URL PMID |
| [14] |
Cao P, Cao DF, Si L, Su XD, Tian LJ, Chang WR, Liu ZF, Zhang XZ, Li M (2020). Structural basis for energy and electron transfer of the photosystem I-IsiA-flavodoxin supercomplex. Nat Plants 6, 167-176.
DOI URL PMID |
| [15] |
Díaz-Quintana A, Leibl W, Bottin H, Sétif P (1998). Electron transfer in photosystem I reaction centers follows a linear pathway in which iron-sulfur cluster FB is the immediate electron donor to soluble ferredoxin. Biochemistry 37, 3429-3439.
URL PMID |
| [16] |
Fristedt R, Williams-Carrier R, Merchant SS, Barkan A (2014). A thylakoid membrane protein harboring a DnaJ-type zinc finger domain is required for photosystem I accumulation in plants. J Biol Chem 289, 30657-30667.
URL PMID |
| [17] | Haehnel W (1984). Photosynthetic electron transport in higher plants. Annu Rev Plant Physiol 35, 659-693. |
| [18] | Harbinson J, Hedley CL (1989). The kinetics of P-700+ reduction in leaves: a novel in situ probe of thylakoid functioning Plant Cell Environ 12, 357-369. |
| [19] |
Harbinson J, Hedley CL (1993). Changes in P-700 oxidation during the early stages of the induction of photosynthesis. Plant Physiol 103, 649-660.
DOI URL PMID |
| [20] | Harbinson J, Woodward FI (1987). The use of light-induced absorbance changes at 820 nm to monitor the oxidation state of P-700 in leaves. Plant Cell Environ 10, 131-140. |
| [21] |
Hiyama T, Ke B (1971). A new photosynthetic pigment, "P430": its possible role as the primary electron acceptor of Photosystem I. Proc Natl Acad Sci USA 68, 1010-1013.
DOI URL PMID |
| [22] |
Iwai M, Takizawa K, Tokutsu R, Okamuro A, Takahashi Y, Minagawa J (2010). Isolation of the elusive supercomplex that drives cyclic electron flow in photosynthesis. Nature 464, 1210-1213.
DOI URL PMID |
| [23] |
Joliot P, Johnson GN (2011). Regulation of cyclic and linear electron flow in higher plants. Proc Natl Acad Sci USA 108, 13317-13322.
DOI URL PMID |
| [24] |
Joliot P, Joliot A (2005). Quantification of cyclic and linear flows in plants. Proc Natl Acad Sci USA 102, 4913-4918.
URL PMID |
| [25] |
Jordan P, Fromme P, Witt HT, Klukas O, Saenger W, Krauß N (2001). Three-dimensional structure of cyanobacterial photosystem I at 2.5 Å resolution. Nature 411, 909-917.
DOI URL PMID |
| [26] |
Kadota K, Furutani R, Makino A, Suzuki Y, Wada S, Miyake C (2019). Oxidation of P700 induces alternative electron flow in photosystem I in wheat leaves. Plants (Basel) 8, 152.
DOI URL |
| [27] |
Ke B (1973). The primary electron acceptor of photosystem I. Biochim Biophys Acta 301, 1-33.
DOI URL PMID |
| [28] | Klughammer C, Schreiber U (1994). An improved method, using saturating light pulses, for the determination of photosystem I quantum yield via P700+-absorbance changes at 830 nm. Planta 192, 261-268. |
| [29] | Klughammer C, Schreiber U (1998). Measuring P700 absorbance changes in the near infrared spectral region with a dual wavelength pulse modulation system. In: Garab G, ed. Photosynthesis: Mechanisms and Effects. Dordrecht: Sprin-ger. pp. 4357-4360. |
| [30] | Klughammer C, Schreiber U (2008). Saturation pulse method for assessment of energy conversion in PS I. PAM Appl Notes 1, 11-14. |
| [31] | Kok B (1957). Light induced absorption changes in photosynthetic organisms. Acta Bot Neerl 6, 316-336. |
| [32] |
Kou JC, Oguchi R, Fan DY, Chow WS (2012). The time course of photoinactivation of photosystem II in leaves revisited. Photosynth Res 113, 157-164.
URL PMID |
| [33] |
Liu J, Yang HY, Lu QT, Wen XG, Chen F, Peng LW, Zhang LX, Lu CM (2012). PsbP-domain protein1, a nuclear-encoded thylakoid lumenal protein, is essential for photosystem I assembly in Arabidopsis. Plant Cell 24, 4992-5006.
URL PMID |
| [34] |
Losciale P, Oguchi R, Hendrickson L, Hope AB, Corelli- Grappadelli L, Chow WS (2008). A rapid, whole- tissue determination of the functional fraction of PSII after photoinhibition of leaves based on flash-induced P700 redox kinetics. Physiol Plant 132, 23-32.
URL PMID |
| [35] |
Maxwell PC, Biggins J (1976). Role of cyclic electron transport in photosynthesis as measured by the photoinduced turnover of P700 in vivo. Biochemistry 15, 3975-3981.
DOI URL PMID |
| [36] |
Meurer J, Meierhoff K, Westhoff P (1996). Isolation of high- chlorophyll-fluorescence mutants of Arabidopsis thaliana and their characterisation by spectroscopy, immunoblotting and northern hybridisation. Planta 198, 385-396.
DOI URL PMID |
| [37] |
Mozzo M, Mantelli M, Passarini F, Caffarri S, Croce R, Bassi R (2010). Functional analysis of Photosystem I light-harve-sting complexes (Lhca) gene products of Chlamydomonas reinhardtii. Biochim Biophys Acta 1797, 212-221.
DOI URL PMID |
| [38] |
Nguyen K, Vaughn M, Frymier P, Bruce BD (2017). In vitro kinetics of P700 + reduction of Thermosynechococcus elongatus trimeric Photosystem I complexes by recombinant cytochrome c6 using a Joliot-type LED spectrophotometer. Photosynth Res 131, 79-91.
DOI URL PMID |
| [39] | Pfündel E, Klughammer C, Schreiber U (2008). Monitoring the effects of reduced PSII antenna size on quantum yields of photosystems I and II using the Dual-PAM-100 measuring system. PAM Appl Notes 1, 21-24. |
| [40] |
Pi X, Tian LR, Dai HE, Qin XC, Cheng LP, Kuang TY, Sui SF, Shen JR (2018). Unique organization of photosystem I- light-harvesting supercomplex revealed by cryo-EM from a red alga. Proc Natl Acad Sci USA 115, 4423-4428.
DOI URL PMID |
| [41] |
Qin XC, Pi X, Wang WD, Han GY, Zhu LX, Liu MM, Cheng LP, Shen JR, Kuang TY, Sui SF (2019). Structure of a green algal photosystem I in complex with a large number of light- harvesting complex I subunits. Nat Plants 5, 263-272.
DOI URL PMID |
| [42] |
Qin XC, Suga M, Kuang TY, Shen JR (2015). Structural basis for energy transfer pathways in the plant PSI-LHCI supercomplex. Science 348, 989-995.
DOI URL PMID |
| [43] |
Schreiber U (2017). Redox changes of ferredoxin, P700, and plastocyanin measured simultaneously in intact leaves. Photosynth Res 134, 343-360.
DOI URL PMID |
| [44] | Schreiber U, Klughammer C, Neubauer C (1988). Measuring P700 absorbance changes around 830 nm with a new type of pulse modulation system. Z Naturforsch C 43, 686-698. |
| [45] |
Sétif P (2001). Ferredoxin and flavodoxin reduction by photosystem I. Biochim Biophys Acta 1507, 161-179.
DOI URL PMID |
| [46] |
Takahashi H, Clowez S, Wollman FA, Vallon O, Rappaport F (2013). Cyclic electron flow is redox-controlled but independent of state transition. Nat Commun 4, 1954.
DOI URL PMID |
| [47] |
Varotto C, Pesaresi P, Meurer J, Oelmüller R, Steiner-Lange S, Salamini F, Leister D (2000). Disruption of the Arabidopsis photosystem I gene psaE1 affects photosynthesis and impairs growth. Plant J 22, 115-124.
DOI URL PMID |
| [48] |
Vassiliev IR, Jung YS, Yang F, Golbeck JH (1998). PsaC subunit of photosystem I is oriented with iron-sulfur cluster FB as the immediate electron donor to ferredoxin and flavodoxin. Biophys J 74, 2029-2035.
DOI URL PMID |
| [49] |
Wang L, Li QW, Zhang AH, Zhou W, Jang R, Yang ZP, Yang HX, Qin XC, Ding SH, Lu QT, Wen XG, Lu CM (2017). The phytol phosphorylation pathway is essential for the biosynthesis of phylloquinone, which is required for photosystem I stability inArabidopsis. Mol Plant 10, 183-196.
DOI URL PMID |
| [50] | Weis E, Ball JT, Berry J (1987). Photosynthetic control of electron transport in leaves of Phaseolus vulgaris: evidence for regulation of photosystem 2 by the proton gradient. In: Biggins J, ed. Progress in Photosynthesis Research. Dordrecht:Springer. pp. 553-556. |
| [51] |
Yang HX, Liu J, Wen XG, Lu CM (2015). Molecular mechanism of photosystem I assembly in oxygenic organisms. Biochim Biophys Acta 1847, 838-848.
DOI URL PMID |
| [52] |
Yang HY, Li P, Zhang AH, Wen XG, Zhang LX, Lu CM (2017). Tetratricopeptide repeat protein Pyg7 is essential for photosystem I assembly by interacting with PsaC in Arabidopsis. Plant J 91, 950-961.
DOI URL PMID |
| [53] |
Zhou W, Lu QT, Li QW, Wang L, Ding SH, Zhang AH, Wen XG, Zhang LX, Lu CM (2017). PPR-SMR protein SOT1 has RNA endonuclease activity. Proc Natl Acad Sci USA 114, E1554-E1563.
DOI URL PMID |
| No related articles found! |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
 首页
首页