
植物学报 ›› 2019, Vol. 54 ›› Issue (5): 596-605.DOI: 10.11983/CBB18261 cstr: 32102.14.CBB18261
杨艺1,王娜1,王奎玲1,刘庆华1,李伟1,郭霄1,马颖杰2,刘庆超1,*( )
)
收稿日期:2018-12-05
接受日期:2019-06-18
出版日期:2019-09-01
发布日期:2020-03-10
通讯作者:
刘庆超
基金资助:
Yi Yang1,Na Wang1,Kuiling Wang1,Qinghua Liu1,Wei Li1,Xiao Guo1,Yingjie Ma2,Qingchao Liu1,*()
Received:2018-12-05
Accepted:2019-06-18
Online:2019-09-01
Published:2020-03-10
Contact:
Qingchao Liu
摘要: 铁线莲属植物在花部形态和结构方面存在较大差异, 遗传背景相对复杂。因此, 在杂交育种前对其进行胚胎学研究具有重要意义。利用石蜡切片技术对大叶铁线莲(Clematis heracleifolia)大小孢子发生及雌雄配子体发育过程进行研究, 结果显示, 大叶铁线莲具雄株和两性花植株。雄花中, 雄配子体发育偶见败育现象; 而两性花中多数花粉发育异常, 形成功能性雌花。正常发育的两性花中, 雄蕊较雌蕊先发育完全。花药4室, 具腺质绒毡层, 偶见变形绒毡层。胞质分裂为同时型, 以四面体型四分体为主, 偶见左右对称型。成熟花药中, 花药壁由纤维状加厚的表皮及药室内壁构成, 花粉粒为2-细胞型, 近球状, 散沟型。子房1室, 内含少量退化胚珠及1个发育正常的胚珠, 倒生, 单珠被, 薄珠心, 蓼型胚囊, 具线形大孢子四分体及双核反足细胞。大叶铁线莲可能处于相对进化的过渡地位。在杂交育种中, 建议以雄花植株作为父本, 两性花植株仅用作母本; 在两性花花芽大小为0.5-0.8 cm时进行去雄处理。
杨艺,王娜,王奎玲,刘庆华,李伟,郭霄,马颖杰,刘庆超. 大叶铁线莲大小孢子发生及雌雄配子体发育. 植物学报, 2019, 54(5): 596-605.
Yi Yang,Na Wang,Kuiling Wang,Qinghua Liu,Wei Li,Xiao Guo,Yingjie Ma,Qingchao Liu. Megasporogenesis, Microsporogenesis and Development of Male and Female Gametophytes of Clematis heracleifolia. Chinese Bulletin of Botany, 2019, 54(5): 596-605.
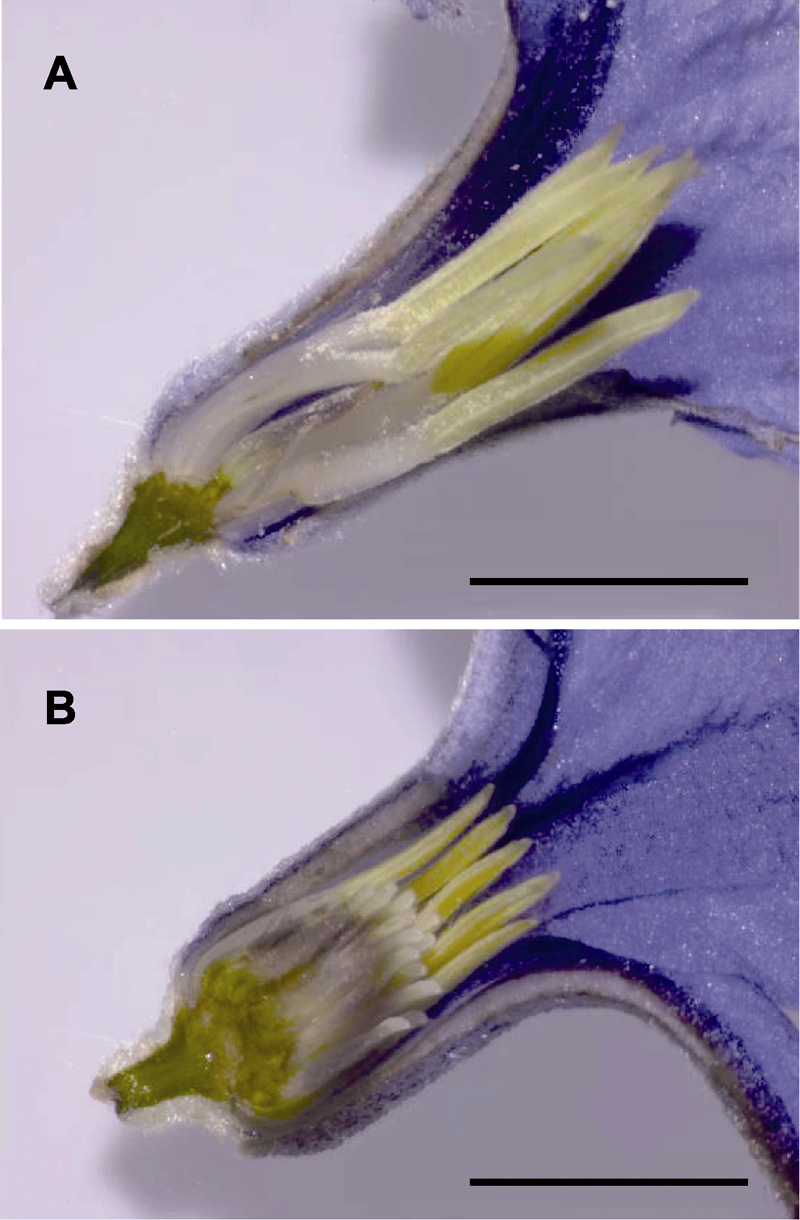
图1 大叶铁线莲花部结构 雄花; (B) 两性花。Bars=5 mm
Figure 1 The floral structure of Clematis heracleifolia Staminate flower; (B) Bisexual flower. Bars=5 mm
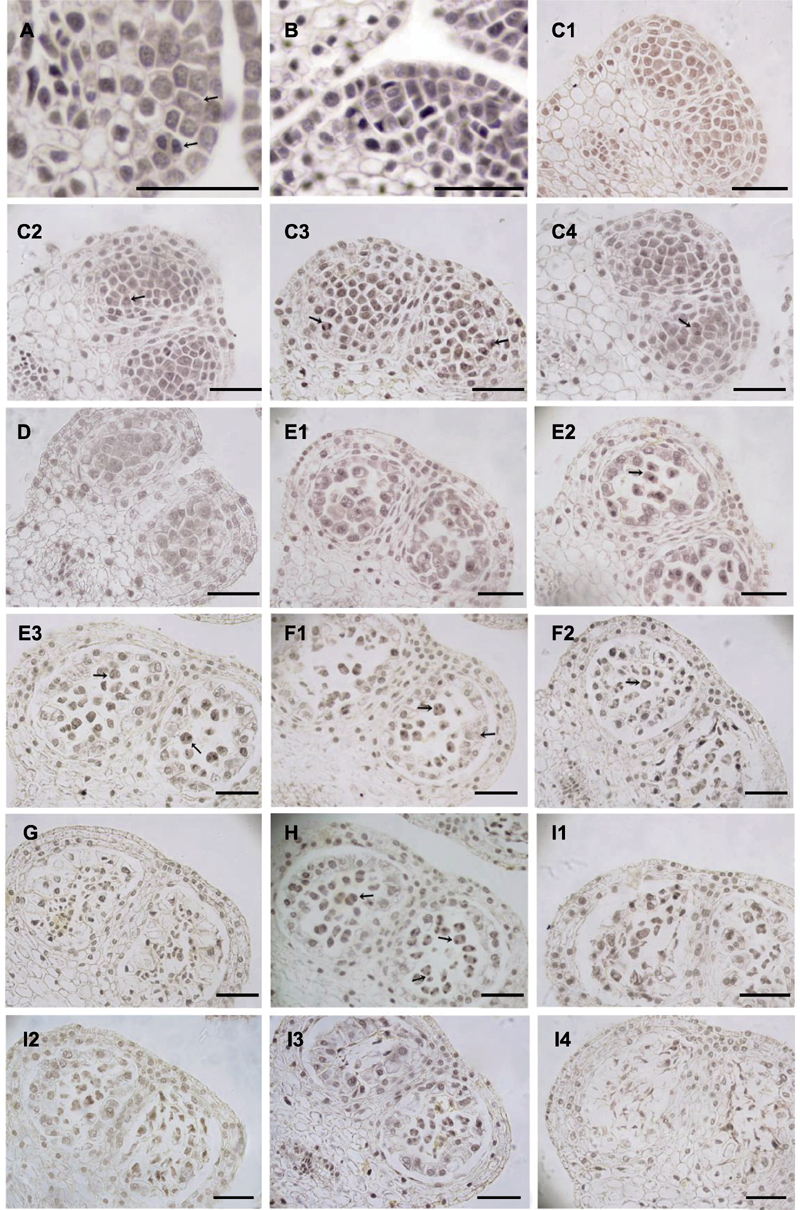
图2 大叶铁线莲雄花小孢子发育 (A) 孢原细胞(箭头所示, 上)及其平周分裂形成初生壁细胞(箭头所示, 上)和初生造孢细胞(箭头所示, 下); (B) 花药壁; (C1) 次生造孢细胞; (C2) 有丝分裂前期(箭头所示); (C3) 有丝分裂中期(箭头所示, 右)和后期(箭头所示, 左); (C4) 有丝分裂末期(箭头所示); (D) 小孢子母细胞; (E1) 减数第1次分裂前期; (E2) 减数第1次分裂中期(箭头所示); (E3) 减数第1次分裂后期(箭头所示, 右)和末期(箭头所示, 左); (F1) 减数第2次分裂后期(箭头所示, 左), 偶见变形绒毡层(箭头所示, 右); (F2) 减数第2次分裂末期, 偶见左右对称型四分体(箭头所示); (G) 四分体解体; (H) 花药发育异步现象(箭头所示); (I1)-(I4) 小孢子败育。Bars=50 μm
Figure 2 Microspore development in staminat e flower of Clematis heracleifolia (A) Archesporial cell (arrow, up), primary parietal cell (arrow, up) and primary sporogenous cell (arrow, down); (B) Anther wall; (C1) A row of secondary sporogenous cells; (C2) Prophase (arrow); (C3) Metaphase (arrow, right) and anaphase (arrow, left); (C4) Telophase (arrow); (D) Microspore mother cells; (E1) Microspore mother cells at prophase of prophase I of meiosis; (E2) Microspore mother cells at metaphase of meiosis I (arrow); (E3) Microspore mother cells at anaphase (arrow, right) and telophase (arrow, left) of meiosis I; (F1) Microspore mother cells at anaphase of meiosis II (arrow, left) and amoeboid tapetum (arrow, right); (F2) Microspore mother cells at telophase of meiosis II (arrow) and symmetrical microspore tetrads; (G) Degrading tetrahedral tetrad; (H) The asynchronous phases; (I1)-(I4) Aborted microspores. Bars=50 μm
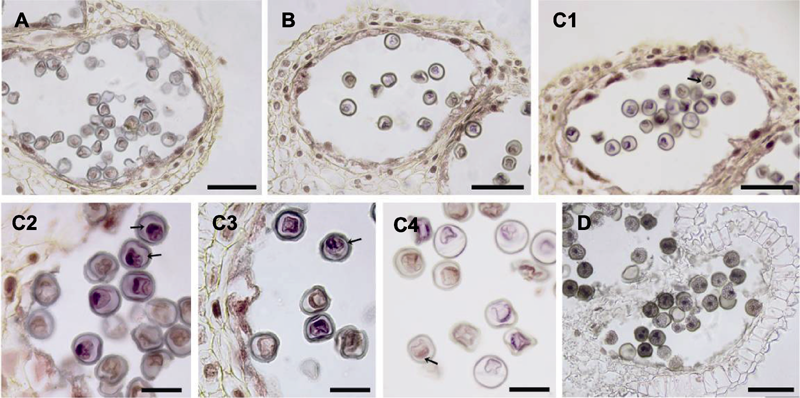
图3 大叶铁线莲雄花雄配子体发育 (A) 单核居中期; (B) 单核靠边期; (C1) 有丝分裂前期(箭头所示); (C2) 有丝分裂中期(箭头所示, 下)和后期(箭头所示, 上); (C3) 有丝分裂后期(箭头所示); (C4) 有丝分裂末期, 形成营养细胞和生殖细胞(箭头所示); (D) 花药开裂, 散粉, 2-细胞花粉粒。(A), (B), (C1), (D) Bars=50 μm; (C2)-(C4) Bars=20 μm
Figure 3 Microgametophyte development in staminate flower of Clematis heracleifolia (A) Microspore at monokaryotic stage; (B) Microspore at monokaryotic side stage; (C1) Microspore at prophase (arrow); (C2) Microspore at metaphase (arrow, down) and anaphase (arrow, up); (C3) Microspore at anaphase (arrow); (C4) Microspore at telophase, vegetative cell and germ cell are formed (arrow); (D) The dehiscent anther, and the 2-celled mature pollen grains. (A), (B), (C1), (D) Bars=50 μm; (C2)-(C4) Bars=20 μm
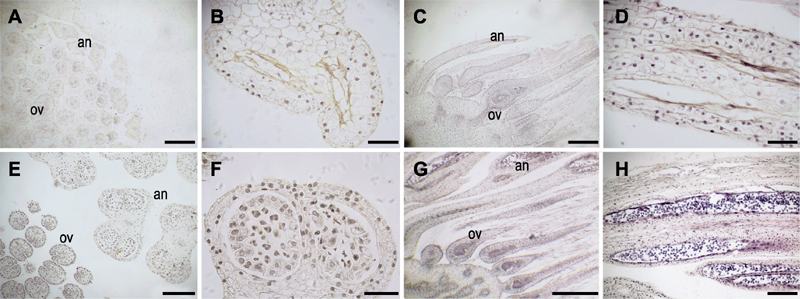
图4 大叶铁线莲两性花小孢子发生及花粉发育 (A)-(D) 小孢子母细胞败育; (E), (F) 部分小孢子母细胞败育; (G), (H) 花粉正常发育。an: 花药; ov: 子房。(A), (C), (G) Bars=500 μm; (B), (F) Bars=50 μm; (D), (E), (H) Bars=200 μm
Figure 4 Microspore and pollen grain development in bisexual flower of Clematis heracleifolia (A)-(D) Aborted microspore mother cells; (E), (F) Some aborted microspore mother cells; (G), (H) Normal pollen grains. an: Anther; ov: Ovary. (A), (C), (G) Bars=500 μm; (B), (F) Bars=50 μm; (D), (E), (H) Bars=200 μm
| Characters | State |
|---|---|
| Shape | Spheroidal |
| Type of aperture | Pantocolpate |
| Diameter (μm) | 16.09±0.88 |
| Spinule height (μm) | 0.46±0.09 |
| Number of spinules (3×3 μm2) | 3.80±1.08 |
表1 大叶铁线莲花粉特性
Table 1 Pollen characteristics of Clematis heracleifolia
| Characters | State |
|---|---|
| Shape | Spheroidal |
| Type of aperture | Pantocolpate |
| Diameter (μm) | 16.09±0.88 |
| Spinule height (μm) | 0.46±0.09 |
| Number of spinules (3×3 μm2) | 3.80±1.08 |
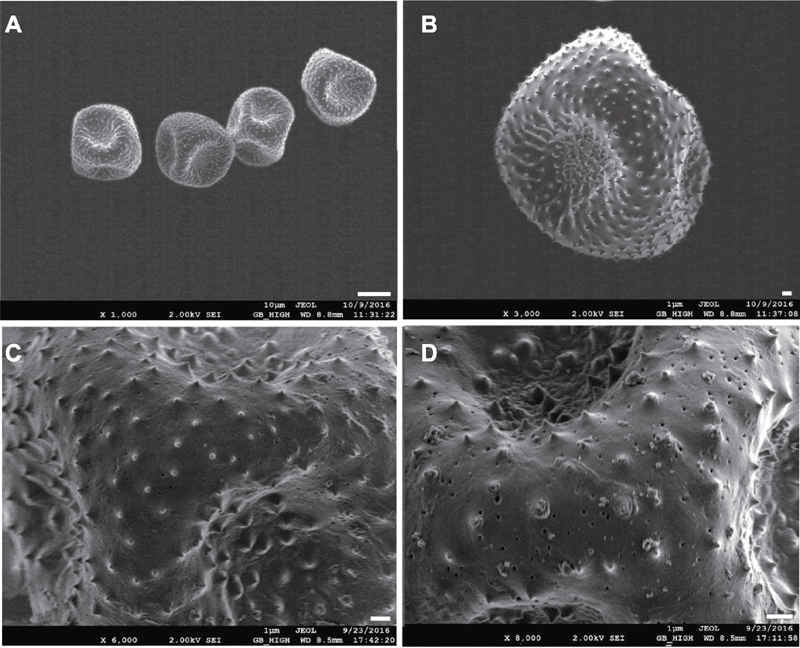
图5 大叶铁线莲花粉扫描电镜观察 (A) Bar=10 μm; (B)-(D) Bars=1 μm
Figure 5 Scanning electron microscopy views of pollen grains of Clematis heracleifolia (A) Bar=10 μm; (B)-(D) Bars=1 μm
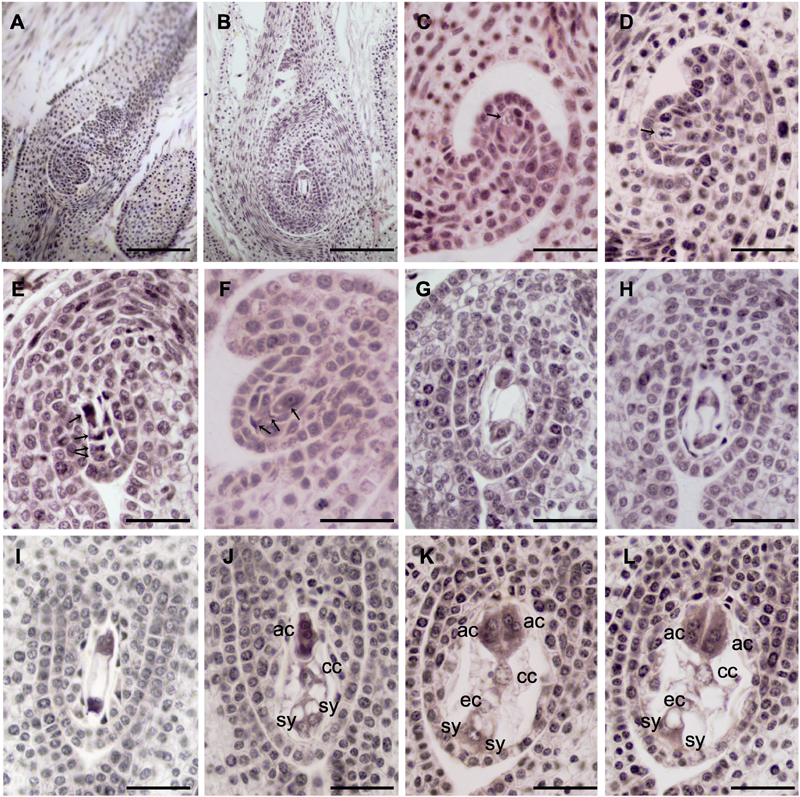
图6 大叶铁线莲两性花大孢子及雌配子体发育 (A), (B) 子房和胚珠; (C) 孢原细胞(箭头所示); (D) 大孢子母细胞(箭头所示); (E) 线形大孢子四分体(箭头所示); (F) 珠孔端3个大孢子退化, 合点端发育形成功能大孢子(箭头所示); (G) 二核胚囊; (H) 四核胚囊; (I) 八核胚囊; (J)-(L) 成熟胚囊7细胞8核。ac: 反足细胞; cc: 中央极核; ec: 卵细胞; sy: 助细胞。(A), (B) Bars=200 μm; (C)-(L) Bars=50 μm
Figure 6 Megaspore and megagametophyte development in bisexual flower of Clematis heracleifolia (A), (B) The ovary and ovule; (C) Archesporial cells (arrow); (D) Megaspore mother cells (arrow); (E) A linear tetrad of megaspores (arrows); (F) The three endostome megaspores were degenerated and the chalazal one was a functional megaspore (arrows); (G) Binucleate embryo sac; (H) Tetranucleate embryo sac; (I) Embryo sac with 8 nuclei; (J)-(L) Mature embryo sac of seven-cell with eight nuclei. ac: Antipodal cell; cc: Central nucleus; ec: Egg cell; sy: Synergids. (A), (B) Bars=200 μm; (C)-(L) Bars=50 μm
| Bud length (cm) | Stamen length (cm) | Male gametophyte |
|---|---|---|
| 0.22-0.40 | 0.15-0.31 | Microsporocyte, meiosis |
| 0.40-0.85 | 0.31-0.51 | Meiosis, microspore |
| 0.85-1.09 | 0.51-0.86 | Microspore, monokaryotic side stage, mitosis |
| 1.09-1.39 | 0.86-1.12 | Mitosis, 2-celled pollen |
| 1.39-2.14 | 1.12-1.32 | 2-celled pollen |
表2 大叶铁线莲雄花中配子体发育与花芽形态的关系
Table 2 Relationship between gametophytes development and flower bud morphology in staminate flower of Clematis heracleifolia
| Bud length (cm) | Stamen length (cm) | Male gametophyte |
|---|---|---|
| 0.22-0.40 | 0.15-0.31 | Microsporocyte, meiosis |
| 0.40-0.85 | 0.31-0.51 | Meiosis, microspore |
| 0.85-1.09 | 0.51-0.86 | Microspore, monokaryotic side stage, mitosis |
| 1.09-1.39 | 0.86-1.12 | Mitosis, 2-celled pollen |
| 1.39-2.14 | 1.12-1.32 | 2-celled pollen |
| Bud length (cm) | Stamen length (cm) | Pistil length (cm) | Male gametophyte | Female gametophyte |
|---|---|---|---|---|
| 0.16-0.36 | 0.10-0.14 | 0.05-0.11 | Microsporocyte, meiosis, microspore | - |
| 0.36-0.50 | 0.14-0.34 | 0.10-0.18 | Microspore monokaryotic side stage, mitosis | Archesporium, megasporocyte |
| 0.50-0.88 | 0.34-0.48 | 0.18-0.21 | Mitosis, 2-celled pollen | Megasporocyte, uninucleate embryo sac |
| 0.88-0.95 | 0.48-0.52 | 0.21-0.30 | 2-celled pollen | Uninucleate embryo sac, 2-nucleate or 4-nucleate embryo sac |
| 0.95-1.42 | 0.52-0.60 | 0.30-0.42 | 2-celled pollen | 4-nucleate or 8-nucleate embryo sac, mature embryo sac |
表3 大叶铁线莲两性花中配子体发育与花芽形态的关系
Table 3 Relationship between gametophytes development and flower bud morphology in bisexual flower of Clematis heracleifolia
| Bud length (cm) | Stamen length (cm) | Pistil length (cm) | Male gametophyte | Female gametophyte |
|---|---|---|---|---|
| 0.16-0.36 | 0.10-0.14 | 0.05-0.11 | Microsporocyte, meiosis, microspore | - |
| 0.36-0.50 | 0.14-0.34 | 0.10-0.18 | Microspore monokaryotic side stage, mitosis | Archesporium, megasporocyte |
| 0.50-0.88 | 0.34-0.48 | 0.18-0.21 | Mitosis, 2-celled pollen | Megasporocyte, uninucleate embryo sac |
| 0.88-0.95 | 0.48-0.52 | 0.21-0.30 | 2-celled pollen | Uninucleate embryo sac, 2-nucleate or 4-nucleate embryo sac |
| 0.95-1.42 | 0.52-0.60 | 0.30-0.42 | 2-celled pollen | 4-nucleate or 8-nucleate embryo sac, mature embryo sac |
| 1 | 管开云, 李志坚, 李景秀, 匡建 (2002). 铁线莲属植物的引种栽培研究初报. 植物分类与资源学报 24, 392-396. |
| 2 | 李杨, 张伟, 赫雪锋, 汤海峰 (2018). 铁线莲属植物的化学成分及药理作用研究进展. 中南药学 16, 355-362. |
| 3 | 刘萍 (2006). 铁筷子(毛茛科) 的形态、结构、发育及系统位置的研究. 硕士论文. 西安: 陕西师范大学. pp. 15-35. |
| 4 | 刘庆超, 王奎玲, 卢婉佩, 刘庆华 (2014). 我国铁线莲属植物资源研究进展. 植物遗传资源学报 15, 483-490, 497. |
| 5 | 全雪丽, 刘继生, 吴松权 (2012). 棉团铁线莲小孢子发生和雄配子体的形成. 辽宁林业科技(1), 9-10, 32. |
| 6 | 宋春慧 (2006). 黄三七属(毛茛科) 植物的形态、结构、发育及系统位置研究. 硕士论文. 西安: 陕西师范大学. pp. 25. |
| 7 | 宋萍 (2007). 金莲花属和驴蹄草属(毛茛科)的比较形态学研究——基于形态、结构、花发育及胚胎学的证据. 硕士论文. 西安: 陕西师范大学. pp. 11-18. |
| 8 | 孙诚, 谢磊, 李良千 (2007). 铁线莲属尾叶铁线莲组(毛茛科)基于形态学证据的分支系统学. 植物学通报 24, 87-98. |
| 9 | 王玖瑞, 刘玲, 刘孟军, 周俊义 (2006). 枣树雄性不育新种质的获得. 园艺学报 33, 374-377. |
| 10 | 王娜, 王奎玲, 刘庆华, 刘庆超 (2017). 七种野生铁线莲属植物核型分析. 草业学报 26(11), 123-130. |
| 11 | 王述彬, 罗向东, 戴亮芳, 钱春桃, 陈劲枫 (2004). 辣(甜)椒细胞质雄性不育系减数分裂和雄配子发生过程. 园艺学报 31, 807-810. |
| 12 | 王文采, 李良千 (2005). 铁线莲属一新分类系统. 植物分类学报 43, 431-488. |
| 13 | 王永周, 古松, 江莎, 任艳萍, 许珂, 李清河 (2008). 臭椿雄蕊的发育和雄配子体的形成. 园艺学报 35, 577-586. |
| 14 | 王自芬 (2008). 毛茛科植物胚珠形态、结构、发育及其系统学意义. 博士论文. 西安: 陕西师范大学. pp. 25. |
| 15 | 杨艺, 王娜, 马颖杰, 王奎玲, 刘庆华, 刘庆超 (2018). 4种铁线莲属植物种子萌发特性. 草业科学 35, 2395-2402. |
| 16 | 杨艺, 王娜, 王奎玲, 刘庆华, 刘庆超 (2019). 铁线莲属4种植物的花芽分化研究. 园艺学报 46, 87-95. |
| 17 | 张阿娟 (2006). 类叶升麻(毛茛科) 形态结构、发育及系统位置的研究. 硕士论文. 西安: 陕西师范大学. pp. 14-27. |
| 18 | 张克中, 赵祥云, 梁励, 黄善武, 张启翔 (2003). 王百合雄性不育突变体小孢子败育的细胞学研究. 北京林业大学学报 25(3), 124-126. |
| 19 | 张敏涛, 张荻, 申晓辉 (2016). ‘东方晨曲’铁线莲小孢子发生和雄配子体发育进程的解剖学研究. 西北植物学报 36, 2213-2219. |
| 20 | 张燕, 黎斌, 李思锋 (2010). 铁线莲属植物分类学及园艺学研究进展. 中国野生植物资源 29(5), 6-10. |
| 21 | 赵桦, 孟华菊, 徐娟 (2011). 七叶树小孢子发生及雄配子体发育研究. 西北植物学报 31, 217-222. |
| 22 | Bhandari NN (1968). Studies in the family Ranunculaceae. X. Embryology of Anemone L. Phytomorphology 18, 487-497. |
| 23 | Xie L, Li LQ (2012). Variation of pollen morphology, and its implications in the phylogeny of Clematis(Ranunculaceae). Plant Syst Evol 298, 1437-1453. |
| 24 | Zhang Q, Hao Q, Guo X, Liu QC, Sun YK, Liu QH, Wang KL (2017). Anther and ovule development in Camellia japonica(Naidong) in relation to winter dormancy: climatic evolution considerations. Flora 233, 127-139. |
| [1] | 李亦超, 陈永生, Denis Sandanov, 罗奥, 吕童, 苏香燕, 刘云鹏, 王庆刚, Viktor Chepinoga, Sergey Dudov, 王伟, 王志恒. 欧亚大陆东部毛茛科植物多样性格局及主导因子[J]. 生物多样性, 2021, 29(5): 561-574. |
| [2] | 张冬瑞, 卜志刚, 陈玲玲, 常缨. 香鳞毛蕨的组织培养和快速繁殖体系构建[J]. 植物学报, 2020, 55(6): 760-767. |
| [3] | 刘魏, 童永鳌, 白洁. 水稻雄配子体发育过程中tRNA片段的生物信息学分析[J]. 植物学报, 2018, 53(5): 625-633. |
| [4] | 张九东, 王琳, 隋洁, 田先华, 任毅. 类叶升麻(毛茛科)自花传粉机制[J]. 植物学报, 2018, 53(2): 212-218. |
| [5] | 彭雄波, 孙蒙祥. 中国科学家在植物受精过程中雌雄配子体信号识别机制研究中取得突破性进展[J]. 植物学报, 2016, 51(2): 145-147. |
| [6] | 王莉;陆彦;金飚*;林明明;陈鹏*. 银杏雌雄配子体发育及胚胎形成的研究进展[J]. 植物学报, 2010, 45(01): 119-127. |
| [7] | 杜红红, 李杨, 李东, 戴绍军, 姜闯道, 石雷. 光照、温度和pH值对小黑桫椤孢子萌发及早期配子体发育的影响[J]. 生物多样性, 2009, 17(2): 182-187. |
| [8] | 钟满香;王瑞珍;路国辉;王英强*. 苞叶姜大小孢子发生和雌雄配子体发育[J]. 植物学报, 2009, 44(06): 694-700. |
| [9] | 陆俊萍;申家恒*;郭德栋;王艳杰. 栽培甜菜花粉发育过程的超微结构[J]. 植物学报, 2009, 44(03): 323-330. |
| [10] | 孟宪利;李勇;刘保东*. 叉蕨科4属5种植物配子体的发育模式及其系统学意义[J]. 植物学报, 2008, 25(03): 298-306. |
| [11] | 杨克珍 叶德. 植物雄配子体发生和发育的遗传调控[J]. 植物学报, 2007, 24(03): 293-301. |
| [12] | 杨维才 石东乔. 植物雌配子体发育研究进展[J]. 植物学报, 2007, 24(03): 302-310. |
| [13] | 王艳杰 申家恒. 柴胡大、小孢子发生及雌、雄配子体发育[J]. 植物学报, 2007, 24(03): 425-432. |
| [14] | 辜天琪;任毅. 黄连属(毛茛科)花的形态发生[J]. 植物学报, 2007, 24(01): 80-86. |
| [15] | 孙诚 谢磊 李良千. 铁线莲属尾叶铁线莲组(毛茛科)基于形态学证据的分支系统学[J]. 植物学报, 2007, 24(01): 87-98. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
 首页
首页