
Chinese Bulletin of Botany ›› 2020, Vol. 55 ›› Issue (3): 287-298.DOI: 10.11983/CBB19105 cstr: 32102.14.CBB19105
• EXPERIMENTAL COMMUNICATIONS • Previous Articles Next Articles
Yang Peng1,Yingjuan Su2,3,*( ),Ting Wang1,*()
),Ting Wang1,*()
Received:2019-06-06
Accepted:2020-03-24
Online:2020-05-01
Published:2020-07-06
Contact:
Yingjuan Su,Ting Wang
Yang Peng, Yingjuan Su, Ting Wang. Intron Loss and Molecular Evolution Rate of rpoC1 in Ferns[J]. Chinese Bulletin of Botany, 2020, 55(3): 287-298.
| Family name | Species | GenBank accession number | Length (bp) |
|---|---|---|---|
| Aspleniaceae | Asplenium pekinense | KY427331 | 152479 |
| As. prolongatum | KY427332 | 151115 | |
| Hymenasplenium unilaterale | KY427350 | 151723 | |
| Athyriaceae | Athyrium anisopterum | NC_035738 | 151284 |
| At. opacum | KY427335 | 150979 | |
| At. sheareri | KY427330 | 151068 | |
| At. sinense | KY427333 | 151319 | |
| Deparia lancea | KY427338 | 151011 | |
| De. pycnosora | KY427339 | 151126 | |
| De. viridifrons | KY427340 | 150939 | |
| Diplazium bellum | KY427343 | 151601 | |
| Di. dilatatum | KY427344 | 151114 | |
| Di. dushanense | KY427345 | 150179 | |
| Di. striatum | KY427346 | 150779 | |
| Di. unilobum | KY427347 | 127840 | |
| Blechnaceae | Austroblechnum melanocaulon | KY427334 | 150202 |
| Woodwardia unigemmata | NC_028543 | 153717 | |
| Cibotiaceae | Cibotium barometz | MH105066 | 166027 |
| Cyatheaceae | Alsophila podophylla | MG262389 | 166151 |
| Al. spinulosa | NC_012818 | 156661 | |
| Cystopteridaceae | Cystopteris chinensis | KY427337 | 151269 |
| Family name | Species | GenBank accession number | Length (bp) |
| Dennstaedtiaceae | Pteridium aquilinum | NC_014348 | 152362 |
| Diplaziopsidaceae | Diplaziopsis cavaleriana | KY427341 | 151934 |
| Dip. javanica | KY427342 | 151496 | |
| Homalosorus pycnocarpos | KY427349 | 152159 | |
| Dryopteridaceae | Cyrtomium devexiscapulae | NC_028542 | 151684 |
| C. falcatum | NC_028705 | 151628 | |
| C. fortunei | MG913607 | 151699 | |
| Dryopteris decipiens | KY427348 | 150987 | |
| Dr. fragrans | KX418656 | 151978 | |
| Equisetaceae | Equisetum arvense | NC_014699 | 133309 |
| E. hyemale | NC_020146 | 131760 | |
| Hymenophyllaceae | Callistopteris apiifolia | MH265125 | 144918 |
| Hymenophyllum holochilum | MH265124 | 142214 | |
| Vandenboschia speciosa | NC_041000 | 146874 | |
| Hypodematiaceae | Hypodematium crenatum | KY427351 | 149794 |
| Lygodiaceae | Lygodium japonicum | NC_022136 | 157260 |
| L. microphyllum | NC_039378 | 158891 | |
| Marattiaceae | Angiopteris angustifolia | NC_026300 | 153596 |
| An. evecta | NC_008829 | 153901 | |
| Marsileaceae | Marsilea crenata | NC_022137 | 151628 |
| Pilularia americana | KY863504 | 153076 | |
| Onocleaceae | Matteuccia struthiopteris | KY427353 | 151003 |
| Onoclea sensibilis | KY427354 | 148395 | |
| Ophioglossaceae | Botrychium ternatum | KM817789 | 139127 |
| Helminthostachys zeylanica | KM817788 | 145120 | |
| Mankyua chejuensis | NC_017006 | 146221 | |
| Osmundaceae | Osmundastrum cinnamomeum | NC_024157 | 142812 |
| Polypodiaceae | Lepisorus clathratus | NC_035739 | 156998 |
| Polypodium glycyrrhiza | KP136832 | 129223 | |
| Psilotaceae | Psilotum nudum | NC_003386 | 138829 |
| Pteridaceae | Adiantum shastense | MG432483 | 150414 |
| Ceratopteris richardii | KM052729 | 148444 | |
| Myriopteris lindheimeri | NC_014592 | 155770 | |
| Rhachidosoraceae | Rhachidosorus consimilis | KY427356 | 152642 |
| Schizaeaceae | Schizaea elegans | KX258660 | 156603 |
| S. pectinata | KX258661 | 156392 | |
| Thelypteridaceae | Ampelopteris prolifera | KY427329 | 151772 |
| Christella appendiculata | NC_035842 | 151571 | |
| Macrothelypteris torresiana | KY427352 | 151130 | |
| Pseudophegopteris aurita | KY427355 | 149917 | |
| Stegnogramma sagittifolia | KY427357 | 151132 | |
| Woodsiaceae | Woodsia macrochlaena | KY427358 | 150987 |
| W. polystichoides | KY427359 | 150685 |
Table 1 Plant materials used in this study and GenBank accession numbers of chloroplast genome sequences
| Family name | Species | GenBank accession number | Length (bp) |
|---|---|---|---|
| Aspleniaceae | Asplenium pekinense | KY427331 | 152479 |
| As. prolongatum | KY427332 | 151115 | |
| Hymenasplenium unilaterale | KY427350 | 151723 | |
| Athyriaceae | Athyrium anisopterum | NC_035738 | 151284 |
| At. opacum | KY427335 | 150979 | |
| At. sheareri | KY427330 | 151068 | |
| At. sinense | KY427333 | 151319 | |
| Deparia lancea | KY427338 | 151011 | |
| De. pycnosora | KY427339 | 151126 | |
| De. viridifrons | KY427340 | 150939 | |
| Diplazium bellum | KY427343 | 151601 | |
| Di. dilatatum | KY427344 | 151114 | |
| Di. dushanense | KY427345 | 150179 | |
| Di. striatum | KY427346 | 150779 | |
| Di. unilobum | KY427347 | 127840 | |
| Blechnaceae | Austroblechnum melanocaulon | KY427334 | 150202 |
| Woodwardia unigemmata | NC_028543 | 153717 | |
| Cibotiaceae | Cibotium barometz | MH105066 | 166027 |
| Cyatheaceae | Alsophila podophylla | MG262389 | 166151 |
| Al. spinulosa | NC_012818 | 156661 | |
| Cystopteridaceae | Cystopteris chinensis | KY427337 | 151269 |
| Family name | Species | GenBank accession number | Length (bp) |
| Dennstaedtiaceae | Pteridium aquilinum | NC_014348 | 152362 |
| Diplaziopsidaceae | Diplaziopsis cavaleriana | KY427341 | 151934 |
| Dip. javanica | KY427342 | 151496 | |
| Homalosorus pycnocarpos | KY427349 | 152159 | |
| Dryopteridaceae | Cyrtomium devexiscapulae | NC_028542 | 151684 |
| C. falcatum | NC_028705 | 151628 | |
| C. fortunei | MG913607 | 151699 | |
| Dryopteris decipiens | KY427348 | 150987 | |
| Dr. fragrans | KX418656 | 151978 | |
| Equisetaceae | Equisetum arvense | NC_014699 | 133309 |
| E. hyemale | NC_020146 | 131760 | |
| Hymenophyllaceae | Callistopteris apiifolia | MH265125 | 144918 |
| Hymenophyllum holochilum | MH265124 | 142214 | |
| Vandenboschia speciosa | NC_041000 | 146874 | |
| Hypodematiaceae | Hypodematium crenatum | KY427351 | 149794 |
| Lygodiaceae | Lygodium japonicum | NC_022136 | 157260 |
| L. microphyllum | NC_039378 | 158891 | |
| Marattiaceae | Angiopteris angustifolia | NC_026300 | 153596 |
| An. evecta | NC_008829 | 153901 | |
| Marsileaceae | Marsilea crenata | NC_022137 | 151628 |
| Pilularia americana | KY863504 | 153076 | |
| Onocleaceae | Matteuccia struthiopteris | KY427353 | 151003 |
| Onoclea sensibilis | KY427354 | 148395 | |
| Ophioglossaceae | Botrychium ternatum | KM817789 | 139127 |
| Helminthostachys zeylanica | KM817788 | 145120 | |
| Mankyua chejuensis | NC_017006 | 146221 | |
| Osmundaceae | Osmundastrum cinnamomeum | NC_024157 | 142812 |
| Polypodiaceae | Lepisorus clathratus | NC_035739 | 156998 |
| Polypodium glycyrrhiza | KP136832 | 129223 | |
| Psilotaceae | Psilotum nudum | NC_003386 | 138829 |
| Pteridaceae | Adiantum shastense | MG432483 | 150414 |
| Ceratopteris richardii | KM052729 | 148444 | |
| Myriopteris lindheimeri | NC_014592 | 155770 | |
| Rhachidosoraceae | Rhachidosorus consimilis | KY427356 | 152642 |
| Schizaeaceae | Schizaea elegans | KX258660 | 156603 |
| S. pectinata | KX258661 | 156392 | |
| Thelypteridaceae | Ampelopteris prolifera | KY427329 | 151772 |
| Christella appendiculata | NC_035842 | 151571 | |
| Macrothelypteris torresiana | KY427352 | 151130 | |
| Pseudophegopteris aurita | KY427355 | 149917 | |
| Stegnogramma sagittifolia | KY427357 | 151132 | |
| Woodsiaceae | Woodsia macrochlaena | KY427358 | 150987 |
| W. polystichoides | KY427359 | 150685 |
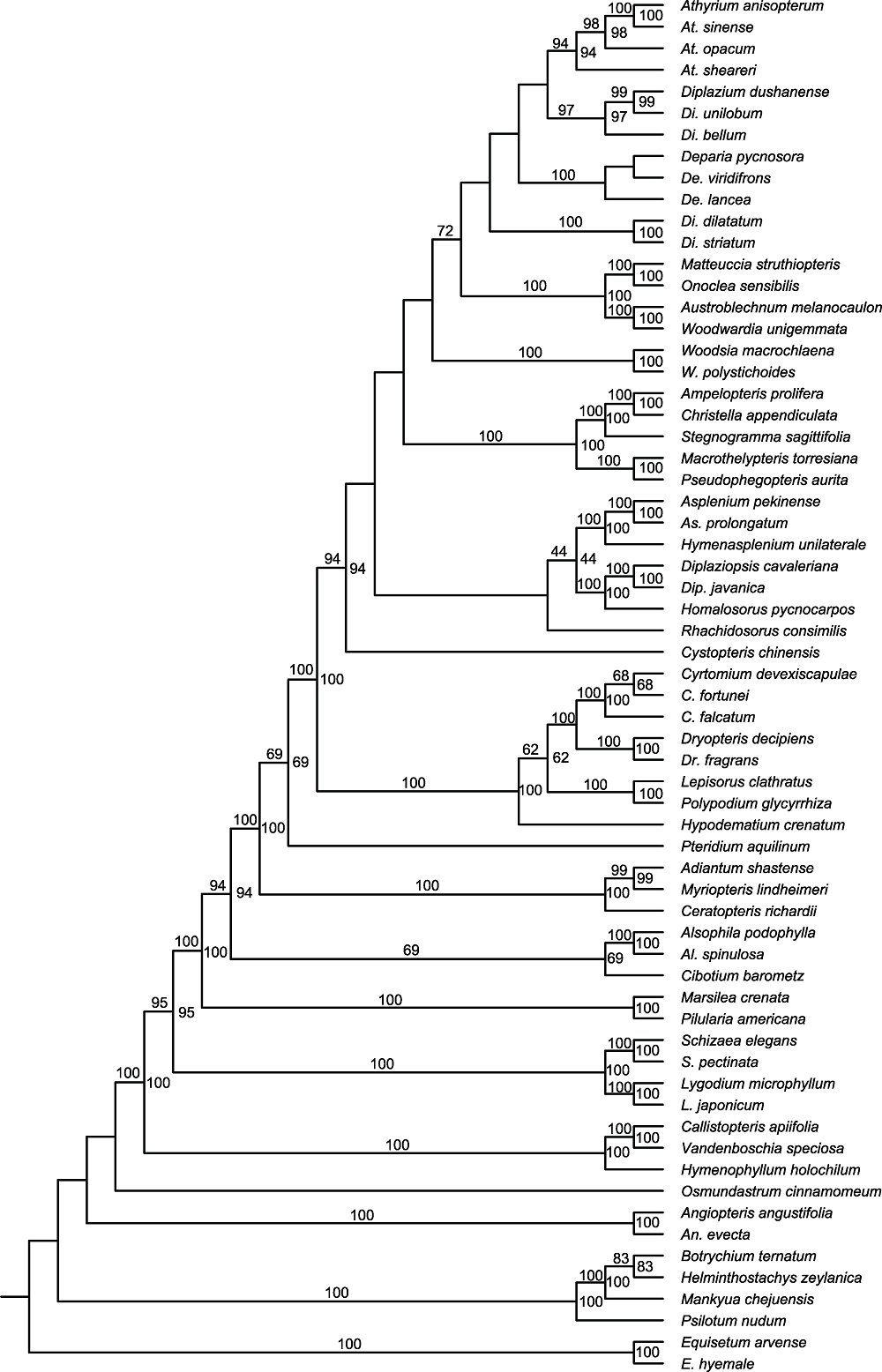
Figure 1 Phylogenetic tree of 64 ferns species
| Et | Ev | trsv/trst | trst | trsv | dN | dS | ω | |
|---|---|---|---|---|---|---|---|---|
| Lygodium | 0.014 | 0.004 | 0.118 | 0.579 | 0.007 | 0.022 | 0.147 | 0.144 |
| Other ferns | 0.022 | 0.005 | 0.096 | 0.088 | 0.010 | 0.045 | 0.201 | 0.256 |
| P value | 1.000 | 1.000 | 1.000 | 0.008 | 0.036 | 0.000 | 0.517 | 1.000 |
| Mann-Whitney | 0.834 | 0.778 | 0.362 | 0.834 | 0.778 | 0.502 | 0.923 | 0.316 |
Table 2 Evolution rate test of rpoC1 coding sequences (CDS) of Lygodium and other ferns
| Et | Ev | trsv/trst | trst | trsv | dN | dS | ω | |
|---|---|---|---|---|---|---|---|---|
| Lygodium | 0.014 | 0.004 | 0.118 | 0.579 | 0.007 | 0.022 | 0.147 | 0.144 |
| Other ferns | 0.022 | 0.005 | 0.096 | 0.088 | 0.010 | 0.045 | 0.201 | 0.256 |
| P value | 1.000 | 1.000 | 1.000 | 0.008 | 0.036 | 0.000 | 0.517 | 1.000 |
| Mann-Whitney | 0.834 | 0.778 | 0.362 | 0.834 | 0.778 | 0.502 | 0.923 | 0.316 |
| Amino acid sites | |
|---|---|
| Positive selection sites | 156, 184, 568 |
| Negative selection sites | 1-3, 6-12, 14-21, 23-26, 28-32, 35-37, 39-42, 44-54, 56, 58-63, 66-74, 76, 79-82, 84, 86-102, 104-107, 109-128, 130-136, 138, 139, 141-144, 147-155, 158-173, 180-181, 183, 186-191, 193-195, 197-198, 200-208, 211, 213-218, 223, 225, 228-230, 232-235, 237, 241-244, 246-247, 249-258, 260-276, 278-291, 294-296, 298-301, 303-315, 318-337, 339-344, 346-368, 370-391, 393-400, 402-411, 413-414, 416-417, 419-434, 436-437, 439-440, 442, 444-445, 447-449, 453-455, 457-480, 482-492, 494-499, 501-526, 528-538, 540-550, 553-556, 558-561, 566-567, 575, 577, 579, 582-583, 586-588, 590-591, 593, 597-600, 603-613, 620, 622, 625-626, 628, 630, 632-635, 639, 642-644, 646, 648-649, 654, 660, 663-668, 671-674, 676, 677, 679-682, 687-689, 694, 697, 710 |
Table 3 Positive and negative selection sites of rpoC1
| Amino acid sites | |
|---|---|
| Positive selection sites | 156, 184, 568 |
| Negative selection sites | 1-3, 6-12, 14-21, 23-26, 28-32, 35-37, 39-42, 44-54, 56, 58-63, 66-74, 76, 79-82, 84, 86-102, 104-107, 109-128, 130-136, 138, 139, 141-144, 147-155, 158-173, 180-181, 183, 186-191, 193-195, 197-198, 200-208, 211, 213-218, 223, 225, 228-230, 232-235, 237, 241-244, 246-247, 249-258, 260-276, 278-291, 294-296, 298-301, 303-315, 318-337, 339-344, 346-368, 370-391, 393-400, 402-411, 413-414, 416-417, 419-434, 436-437, 439-440, 442, 444-445, 447-449, 453-455, 457-480, 482-492, 494-499, 501-526, 528-538, 540-550, 553-556, 558-561, 566-567, 575, 577, 579, 582-583, 586-588, 590-591, 593, 597-600, 603-613, 620, 622, 625-626, 628, 630, 632-635, 639, 642-644, 646, 648-649, 654, 660, 663-668, 671-674, 676, 677, 679-682, 687-689, 694, 697, 710 |
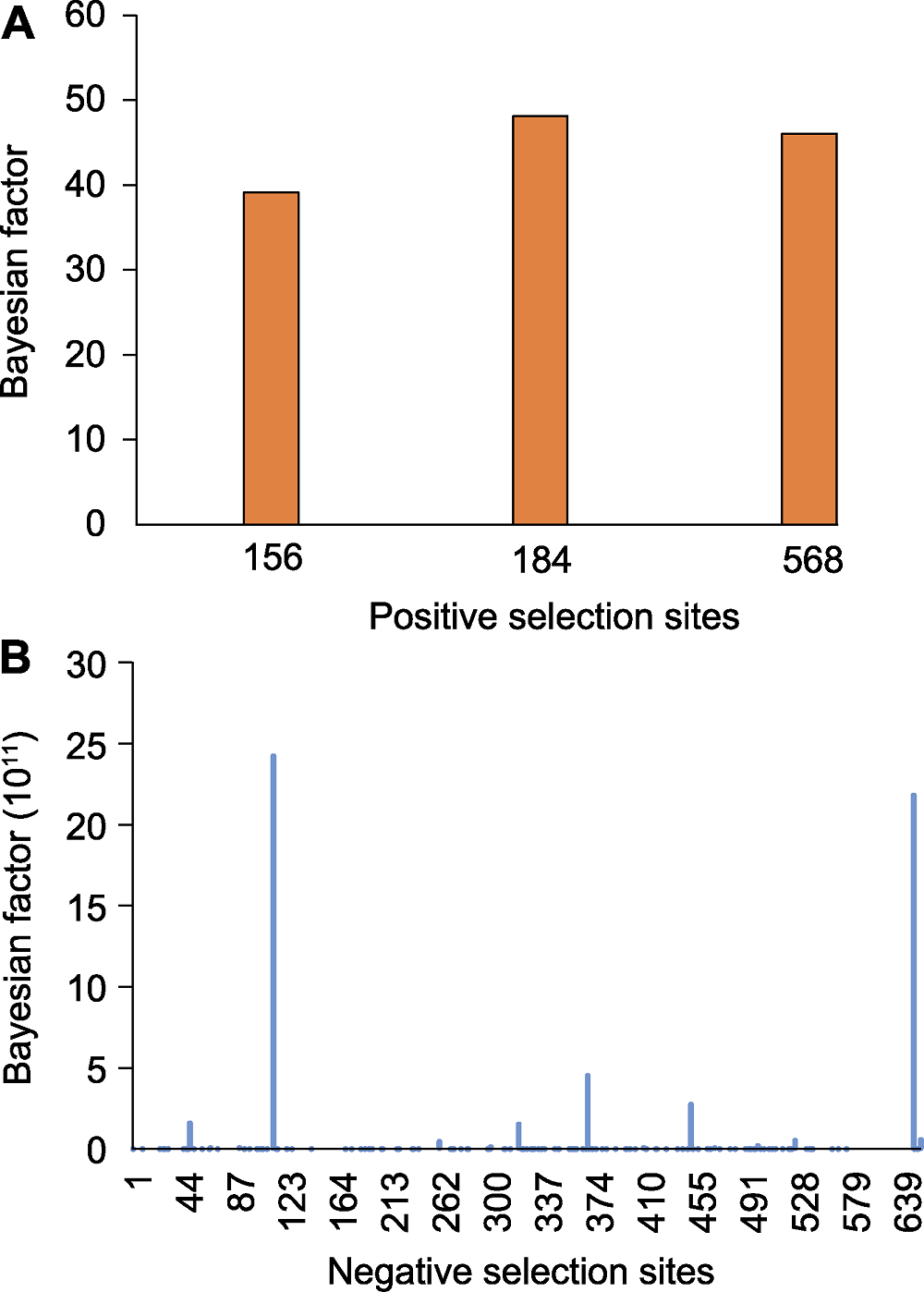
Figure 2 Positive (A) and negative (B) selection sites of rpoC1 amino acid residues
| Model | Np | ? | Parameter estimate | Positive selection sites | |
|---|---|---|---|---|---|
| Branch model | M0 | 128 | -33984.0181 | ω=0.18862 | Not allowed |
| MA | 129 | -33971.0708 | ω1=0.19344, ω2=0.07154 | Not allowed | |
| Site model | Model 1a (M1a) | 129 | -33025.1095 | P0=0.77581, ω0=0.09604 | Not allowed |
| P1=0.22419, ω1=1.00000 | |||||
| Model 2a (M2a) | 131 | -33003.8671 | P0=0.77323, ω0=0.09782 | 687P**, 697S** | |
| P1=0.20583, ω1=1.00000 | 692S**, 700A* | ||||
| P2=0.02094, ω2=2.53148 | |||||
| Model 3 (M3) | 132 | -32770.6265 | P0=0.45915, ω0=0.02383 | None | |
| P1=0.39593, ω1=0.23422 | |||||
| P2=0.14492, ω2=0.91531 | |||||
| Model 7 (M7) | 129 | -32768.3591 | P=0.36168, q=1.11178 | Not allowed | |
| Model 8 (M8) | 131 | -32715.0671 | P0=0.95310, P=0.46388 | 578V*, 579Y*, 686S* | |
| q=2.02806, P1=0.04690 | 687P**, 689T*, 692S** | ||||
| ω=1.62513 | 693I*, 696T*, 697S**, 700A* | ||||
| Model 8a (M8a) | 130 | -32735.2130 | P0=0.91188, P=0.48169 | None | |
| q=2.38635, P1=0.08812 | |||||
| ω=1.00000 | |||||
| Branch-site model | Ma0 | 130 | -33025.1095 | P0=0.77581, P1=0.22419 | Not allowed |
| P2a+P2b=0.00000, ω2=1.00000 | |||||
| Ma1 | 131 | -33025.1095 | P0=0.77581, P1=0.22419 | None | |
| P2a+P2b=0.00000, ω2=1.00000 |
Table 4 Parameters estimates and log-likelihood values of the rpoC1 gene under different models
| Model | Np | ? | Parameter estimate | Positive selection sites | |
|---|---|---|---|---|---|
| Branch model | M0 | 128 | -33984.0181 | ω=0.18862 | Not allowed |
| MA | 129 | -33971.0708 | ω1=0.19344, ω2=0.07154 | Not allowed | |
| Site model | Model 1a (M1a) | 129 | -33025.1095 | P0=0.77581, ω0=0.09604 | Not allowed |
| P1=0.22419, ω1=1.00000 | |||||
| Model 2a (M2a) | 131 | -33003.8671 | P0=0.77323, ω0=0.09782 | 687P**, 697S** | |
| P1=0.20583, ω1=1.00000 | 692S**, 700A* | ||||
| P2=0.02094, ω2=2.53148 | |||||
| Model 3 (M3) | 132 | -32770.6265 | P0=0.45915, ω0=0.02383 | None | |
| P1=0.39593, ω1=0.23422 | |||||
| P2=0.14492, ω2=0.91531 | |||||
| Model 7 (M7) | 129 | -32768.3591 | P=0.36168, q=1.11178 | Not allowed | |
| Model 8 (M8) | 131 | -32715.0671 | P0=0.95310, P=0.46388 | 578V*, 579Y*, 686S* | |
| q=2.02806, P1=0.04690 | 687P**, 689T*, 692S** | ||||
| ω=1.62513 | 693I*, 696T*, 697S**, 700A* | ||||
| Model 8a (M8a) | 130 | -32735.2130 | P0=0.91188, P=0.48169 | None | |
| q=2.38635, P1=0.08812 | |||||
| ω=1.00000 | |||||
| Branch-site model | Ma0 | 130 | -33025.1095 | P0=0.77581, P1=0.22419 | Not allowed |
| P2a+P2b=0.00000, ω2=1.00000 | |||||
| Ma1 | 131 | -33025.1095 | P0=0.77581, P1=0.22419 | None | |
| P2a+P2b=0.00000, ω2=1.00000 |
| Model comparison | Df. | 2Δl | P value |
|---|---|---|---|
| M0 & MA | 1 | 25.8947 | 3.606×10-7 |
| M1a & M2a | 2 | 42.4848 | 5.950×10-10 |
| M0 & M3 | 4 | 2426.7832 | 0 |
| M7 & M8 | 2 | 106.5838 | 0 |
| M8a & M8 | 1 | 40.2917 | 2.187×10-10 |
| M0 & MF | 125 | 169.4740 | 4.998×10-3 |
Table 5 Likelihood ratio test statistic (2Δ?) for different models in PAML4.9
| Model comparison | Df. | 2Δl | P value |
|---|---|---|---|
| M0 & MA | 1 | 25.8947 | 3.606×10-7 |
| M1a & M2a | 2 | 42.4848 | 5.950×10-10 |
| M0 & M3 | 4 | 2426.7832 | 0 |
| M7 & M8 | 2 | 106.5838 | 0 |
| M8a & M8 | 1 | 40.2917 | 2.187×10-10 |
| M0 & MF | 125 | 169.4740 | 4.998×10-3 |
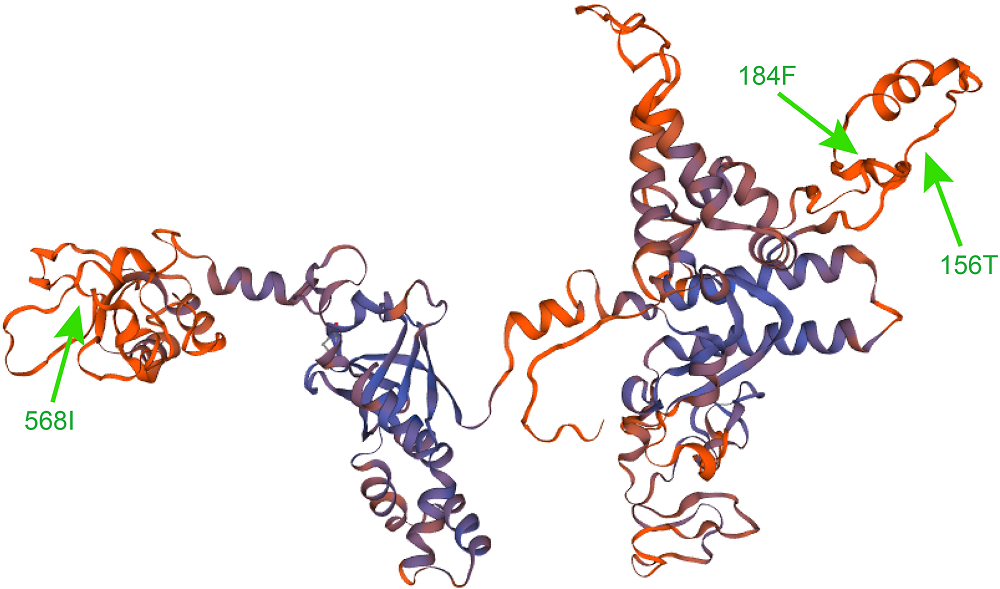
Figure 3 Three-dimensional structure of RPOC1 protein in Lygodium japonicum
| [1] | 中国科学院中国植物志编辑委员会 (1959). 中国植物志, Vol.2. 北京: 科学出版社. pp. 106. |
| [2] | Akaike H (1974). A new look at the statistical model identification. IEEE Trans Autom Control 19, 716-723. |
| [3] |
Darriba D, Taboada GL, Doallo R, Posada D (2012). jModelTest 2: more models, new heuristics and parallel computing. Nat Methods 9, 772.
URL PMID |
| [4] | Downie SR, Katz-Downie DS, Rogers EJ, Zujewski HL, Small E (1998). Multiple independent losses of the plastid rpoC1 intron in Medicago (Fabaceae) as inferred from phylogenetic analyses of nuclear ribosomal DNA internal transcribed spacer sequences. Can J Bot 76, 791-803. |
| [5] | Downie SR, Llanas E, Katz-Downie DS (1996). Multiple independent losses of the rpoC1 intron in angiosperm chloroplast DNA’s. Syst Bot 21, 135-151. |
| [6] |
Dubuisson JY (1997). rbcL sequences: a promising tool for the molecular systematics of the fern genus Trichomanes (Hymenophyllaceae)? Mol Phylogenet Evol 8, 128-138.
DOI URL |
| [7] |
Gao L, Wang B, Wang ZW, Zhou Y, Su YJ, Wang T (2013). Plastome sequences of Lygodium japonicum and Marsilea crenata reveal the genome organization transformation from basal ferns to core Leptosporangiates. Genome Biol Evol 5, 1403-1407.
DOI URL PMID |
| [8] | Guillon JM (2004). Phylogeny of Horsetails (Equisetum) based on the chloroplast rps4 gene and adjacent noncoding sequences. Syst Bot 29, 251-259. |
| [9] |
Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O (2010). New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol 59, 307-321.
URL PMID |
| [10] | Hall TA (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/ 98/NT. Nucl Acid Symp Ser 41, 95-98. |
| [11] | Hansen AK, Gilbert LE, Simpson BB, Downie SR, Cervi AC, Jansen RK (2006). Phylogenetic relationships and chromosome number evolution in Passiflora. Syst Bot 31, 138-150. |
| [12] | He L, Qian J, Li XW, Sun ZY, Xu XL, Chen SL (2017). Complete chloroplast genome of medicinal plant Lonicera japonica: genome rearrangement, intron gain and loss, and implications for phylogenetic studies. Molecules 22, 249. |
| [13] |
Hong XW, Zhang YP, Chu YW, Gao HF, Jiang ZG, Xiong SD (2008). Complete sequence determination and phylogenetic analysis of FKN among seven higher primates including homonids and old world monkeys. Hereditas 30, 595-601.
URL PMID |
| [14] |
Katayama H, Ogihara Y (1993). Structural alterations of the chloroplast genome found in grasses are not common in monocots. Curr Genet 23, 160-165.
DOI URL PMID |
| [15] |
Kim HT, Chung MG, Kim KJ (2014). Chloroplast genome evolution in early diverged leptosporangiate ferns. Mol Cells 37, 372-382.
URL PMID |
| [16] | Korall P, Conant DS, Schneider H, Ueda K, Nishida H, Pryer KM (2006). On the phylogenetic position of Cystodium: it’s not a tree fern—it’s a polypod! Am Fern J 96, 45-53. |
| [17] | Lovis JD (1978). Evolutionary patterns and processes in ferns. Adv Bot Res 4, 229-415. |
| [18] |
Morgan JT, Fink GR, Bartel DP (2019). Excised linear introns regulate growth in yeast. Nature 565, 606-611.
DOI URL PMID |
| [19] |
Newcomb RD, Campbell PM, Ollis DL, Cheah E, Russell RJ, Oakshott JG (1997). A single amino acid substitution converts a carboxylesterase to an organophosphorus hydrolase and confers insecticide resistance on a blowfly. Proc Natl Acad Sci USA 94, 7464-7468.
DOI URL PMID |
| [20] |
Nielsen R, Yang Z (1998). Likelihood models for detecting positively selected amino acid sites and applications to the HIV-1 envelope gene. Genetics 148, 929-936.
URL PMID |
| [21] |
Parenteau J, Maignon L, Berthoumieux M, Catala M, Gagnon V, Abou Elela S (2019). Introns are mediators of cell response to starvation. Nature 565, 612-617.
DOI URL PMID |
| [22] |
Perutz MF (1983). Species adaptation in a protein molecule. Mol Biol Evol 1, 1-28.
URL PMID |
| [23] |
Pond SLK, Frost SDW, Muse SV (2005). HyPhy: hypothesis testing using phylogenies. Bioinformatics 21, 676-679.
URL PMID |
| [24] |
Rothwell GW (1987). Complex paleozoic filicales in the evolutionary radiation of ferns. Am J Bot 74, 458-461.
DOI URL |
| [25] | Schuettpelz E, Pryer KM (2007). Fern phylogeny inferred from 400 leptosporangiate species and three plastid genes. Taxon 56, 1037-1050. |
| [26] | Smith AR, Pryer KM, Schuettpelz E, Korall P, Schneider H, Wolf PG (2006). A classification for extant ferns. Taxon 55, 705-731. |
| [27] | Taylor TN, Taylor EL, Krings M (2009). Paleobotany: the Biology and Evolution of Fossil Plants, 2nd edn. Amsterdam: Academic Press. pp. 1-1252. |
| [28] | The Pteridophyte Phylogeny Group (2016). A community-derived classification for extant lycophytes and ferns. J Syst Evol 54, 563-603. |
| [29] | Thiede J, Schmidt SA, Rudolph B (2007). Phylogenetic implication of the chloroplast rpoC1 intron loss in the Aizoaceae (Caryophyllales). Biochem Syst Ecol 35, 372-380. |
| [30] |
Wallace RS, Cota JH (1996). An intron loss in the chloroplast gene rpoC1 supports a monophyletic origin for the subfamily Cactoideae of the Cactaceae. Curr Genet 29, 275-281.
DOI URL PMID |
| [31] |
Weng ML, Blazier JC, Govindu M, Jansen RK (2014). Reconstruction of the ancestral plastid genome in Geraniaceae reveals a correlation between genome rearrangements, repeats, and nucleotide substitution rates. Mol Biol Evol 31, 645-659.
URL PMID |
| [32] |
Yang ZH (1997). PAML: a program package for phylogenetic analysis by maximum likelihood. CABIOS 13, 555-556.
URL PMID |
| [33] |
Yang ZH (1998). Likelihood ratio tests for detecting positive selection and application to primate lysozyme evolution. Mol Biol Evol 15, 568-573.
DOI URL PMID |
| [34] |
Yang ZH (2005). Bayes empirical Bayes inference of amino acid sites under positive selection. Mol Biol Evol 22, 1107-1118.
URL PMID |
| [35] |
Yang ZH (2007). PAML 4: phylogenetic analysis by maximum likelihood. Mol Biol Evol 24, 1586-1591.
DOI URL PMID |
| [36] |
Yang ZH, Nielsen R (2002). Codon-substitution models for detecting molecular adaptation at individual sites along specific lineages. Mol Biol Evol 19, 908-917.
DOI URL PMID |
| [37] |
Yang ZH, Swanson WJ, Vacquier VD (2000). Maximum- likelihood analysis of molecular adaptation in abalone sperm lysin reveals variable selective pressures among lineages and sites. Mol Biol Evol 17, 1446-1455.
DOI URL PMID |
| [38] |
Zhang JZ, Nielsen R, Yang ZH (2005). Evaluation of an improved branch-site likelihood method for detecting positive selection at the molecular level. Mol Biol Evol 22, 2472-2479.
DOI URL PMID |
| [39] | Zuckerkandl E, Pauling LE (1962). Molecular disease, evolution, and genic heterogeneity. In: Kasha M, Pullman B, eds. Horizons in Biochemistry. New York: Academic Press. pp. 189-225. |
| [1] | WEI Ze-Xiu, LIANG Yin-Li, YAMADA Satoshi, ZENG Xing-Quan, ZHOU Mao-Juan, HUANG Mao-Lin, WU Yan. RELATION OF SOIL MICROBIAL DIVERSITY TO TOMATO YIELD AND QUALITY UNDER DIFFERENT SOIL WATER CONDITIONS AND FERTILIZATIONS [J]. Chin J Plant Ecol, 2009, 33(3): 580-586. |
| [2] | Zhiying Sun;Jiqing Zheng;Fazeng Li* . Taxonomic Position of Arabidopsis (Brassicaceae): Evidence from Seed-coat Micromorphology and Chloroplast DNA trnL-F Sequencing [J]. Chinese Bulletin of Botany, 2008, 25(05): 565-573. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||
 Home
Home