植物miRNA响应非生物胁迫研究进展
周文杰, 张文瀚, 贾玮, 许自成, 黄五星
植物学报
2024, 59 ( 5):
810-833.
DOI: 10.11983/CBB24020
干旱、极端温度、盐和重金属等非生物胁迫导致植物产量和品质下降。miRNA是一类长约20-24个核苷酸的内源性非编码小分子RNA, 通过形成miRNA介导的沉默复合物(RISC)剪切靶mRNA并抑制靶基因的翻译, 在转录后水平负调控真核生物基因表达。高通量测序技术的快速发展使得植物物种中大量响应非生物胁迫的miRNA得到鉴定和表征。非生物胁迫下植物miRNA与其靶基因结合, 构成了控制各种生命活动的大型基因调控网络, 包括生长发育、营养吸收与分配、信号转导与氧化应激, 从而提高植物的抗逆性。深入理解miRNA的功能及其调控机制对于通过基因工程进行作物改良和抗逆育种至关重要。该文综述了近年来miRNA的生物合成及其作用机制研究进展, 重点探讨了参与调控植物响应非生物胁迫miRNA的鉴定及功能, 并展望了该领域可能的研究方向。
View image in article
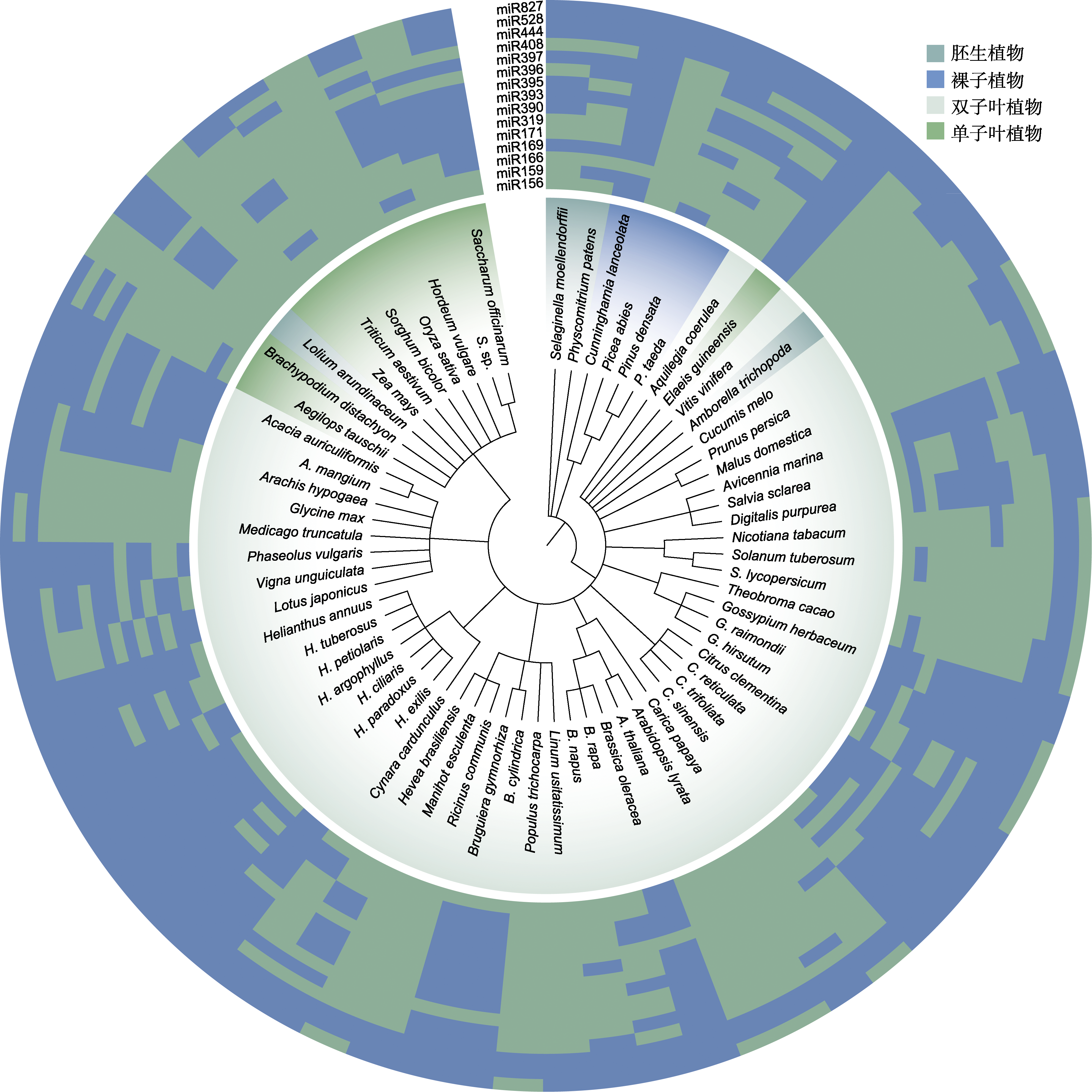
图2
响应植物非生物胁迫miRNAs在不同植物物种中的分布
含有miRNA的植物物种按照NCBI分类进化树进行排列, 热图基质中的绿色元素表示该miRNA在该物种中存在, 蓝色元素表示该miRNA在该物种中不存在。
正文中引用本图/表的段落
多种植物中含有大多数的miRNA家族, miRNA在物种中的进化具有保守性(图2)。miRNA的合成需要DCL1蛋白和HYL1蛋白的加工(Han et al., 2004; Achkar et al., 2018; Wei et al., 2021b)。HYL1-DCL1的互作关系可以追溯到第一批陆地植物中, 来自祖先开花植物的HYL1蛋白可以驱动产生接近原生的miRNA, 以恢复拟南芥HYL1敲除型植株HYL1蛋白的合成(Silvestro et al., 2021; Jia et al., 2021)。参与miRNA发生过程的其它家族蛋白在物种间也表现出强烈的保守性(Murphy et al., 2008)。研究发现, 高等植物中几乎所有的miRNA生物发生因子也存在于苔藓植物中, 这表明陆地植物进化早期就出现了miRNA (Pietrykowska et al., 2022; Velandia-Huerto et al., 2022)。因此, miRNA家族在植物物种进化中持续存在, 是植物基因组中最保守的元素之一。这表明某些miRNA的功能也可能保守, 在多种植物中发挥类似的作用。目前, 已鉴定出许多响应非生物胁迫的miRNA, 涵盖植物应对非生物胁迫的多种途径, 主要包括调节生长发育、营养吸收与分配和信号转导及减少氧化应激损伤(表2; 图3)。
miR156/SPL模块可提高植物对非生物胁迫的耐受性(Wang et al., 2023c; Yuan et al., 2023).利用RLM-5′ RACE技术分析苹果miR156与MdSPL13的关系, 发现MdSPL13的3′UTR区域存在miR156靶标, 证实苹果miR156的靶点为MdSPL13.进一步验证表明, MdSPL13靶向MdWRKY100基因启动子, 因此在苹果中miR156/SPL模块通过靶向调控MdWRKY100调节其耐盐性(Ma et al., 2021).紫花苜蓿(Medicago sativa) miR156过表达植株与野生型相比, 地上部生长更加茂盛, 产量及品质更高, 在盐胁迫下过表达植株中盐含量显著低于野生型, 表现出更强的耐盐性(Wang et al., 2021a).对SPL13 RNAi植株施加干旱胁迫后, 发现叶片中差异表达的基因多集中在光反应系统中, 包括光系统I、II和电子传递链, 说明miR156在叶片中可靶向SPL13调节植物的光合作用, 以抵御干旱胁迫(Feyissa et al., 2020).miR156还靶向SPL2、SPL10、SPL16、SPL18和SPL23等SPL家族基因, 参与植物根系发育、营养生长及生长阶段的转变等重要生命活动(Barrera-Rojas et al., 2020; Su et al., 2023; Zhou and Yarra, 2023; Wang et al., 2023b; Wei et al., 2024). ... PmiREN2.0: from data annotation to functional exploration of plant microRNAs 1 2022b ... miRNA是一类长约20-24个核苷酸的内源性非编码小分子RNA, 通过剪切靶mRNA和抑制靶基因的翻译, 在转录后水平负调控真核生物基因的表达.miRNA于1993年从线虫(Caenorhabditis elegans)中被发现(Lee et al., 1993; Wightman et al., 1993; Fire et al., 1998), 随后于2002年在模式植物拟南芥(Arabidopsis thaliana)中被鉴定发现(Reinhart et al., 2002).随着高通量测序技术和生物学分析手段的发展, 研究人员陆续从其它物种中鉴定出多个miRNA.目前miRBase (http://mirbase.org/)以及PmiREN2.0 (https://pmiren.com/)等miRNA数据库中涵盖了近300个物种的miRNA序列信息(Kozomara et al., 2019; Guo et al., 2022b). ... Characterization of phosphorus-regulated miR399 and miR827 and their isomirs in barley under phosphorus- sufficient and phosphorus-deficient conditions 1 2013 ... miR827可靶向含SPX结构域的基因, 但不同物种中靶基因的进化存在差异.拟南芥miR827靶向氮限制适应基因(NLA), 水稻miR827靶向磷酸盐转运蛋白基因(PHT5), 调节磷酸盐的运输和储存(Hsieh et al., 2009; Lin et al., 2010, 2018; Wang et al., 2015; Liu et al., 2016).缺磷条件下miR827受到强烈诱导以响应磷酸盐的调控过程(Hackenberg et al., 2013).烟草miR827在缺磷处理下表达上调, 表现出对磷缺乏的正响应, 与拟南芥和水稻等表达模式一致(顾冕等, 2016).研究发现, 低氮处理下拟南芥miR827表达量增加, 靶基因NLA转录本无显著变化, 但蛋白表达丰度随缺氮时间的延长显著下降, 说明miR827通过抑制靶基因NLA翻译调控植物的氮分配过程(刘文文, 2016). ... 1 2016 ... 非生物胁迫可上调或下调miRNA在植物中的表达水平.在150 mmol·L-1 NaCl胁迫下, 苹果miR156a的表达随胁迫时间的推移而降低(Ma et al., 2021).番茄和拟南芥中的miR169受高温胁迫诱导上调表达(Rao et al., 2022).干旱胁迫显著上调水稻旗叶和根系中miR408及其前体pre-miR408的丰度(Balyan et al., 2023).但相同miRNA在不同物种中的表达也存在差异.干旱胁迫诱导水稻(Lu et al., 2018)和拟南芥(Liu et al., 2008) miR393表达上调, 但miR393在高粱(Sorghum bicolor) (Hamza et al., 2016)和野生桔梗(Ipomoea campanulata) (Ghorecha et al., 2014)中的表达却受到干旱抑制, 大麦(Hordeum vulgare) miR393过表达植株表现为对干旱更加敏感(Yuan et al., 2019b; Jiang et al., 2022a).盐胁迫下, miR396在西伯利亚翦股颖(Agrostis stolonifera) (Yuan et al., 2019a)、番茄和烟草(Nicotiana tabacum) (Chen et al., 2015)中表达量增高, 在水稻(Gao et al., 2010)、旱柳(Salix matsudana) (Zhou et al., 2012)和拟南芥(Pegler et al., 2021a)中表达量降低.在同种非生物胁迫下, 相同miRNA在不同植物物种中表达模式相反, 表明miRNA在不同植物响应相同非生物胁迫时可能发挥不同的作用.未来仍需要对非生物胁迫下植物miRNA进行大规模鉴定及表达模式分析, 从而进一步阐明miRNA调控网络在植物体内的运行机制. ... Function and regulation of microRNA171 in plant stem cell homeostasis and developmental programing 1 2022 ... miR171启动子上存在光、非生物胁迫以及与胁迫相关的外源激素信号响应元件, 在转录后水平调控SCL转录因子表达(Li et al., 2021; Pei et al., 2023).拟南芥miR171家族受盐、温度和干旱等非生物胁迫信号调控, 过表达株系生物量和抗氧化酶活性均高于野生型, 表现出更强的抗性(Liu et al., 2008; Sun et al., 2022; Han and Zhou, 2022).光信号诱导miR171表达, miR171的表达丰度在光照期增高, 黑暗期降低(Sire et al., 2009).生长素诱导miR171通过靶向PyrSCL6和PyrSCL22负调控生长素信号级联, 促进植物生长发育(Jiang et al., 2018); Sly-miR171沉默株系及靶基因SlGRAS24过表达株系中许多赤霉素和生长素相关基因下调, 表明番茄miR171可调节赤霉素以及生长素信号转导(Huang et al., 2017).水稻miR171/SCL6模块调控类黄酮生物合成, 干旱胁迫下过表达植株叶片温度较低, 茎秆高度及谷物产量更高, 抗旱性增强(Um et al., 2022). ... The Arabidopsis double-stranded RNA-binding protein HYL1 plays a role in microRNA-mediated gene regulation 1 2004 ... 多种植物中含有大多数的miRNA家族, miRNA在物种中的进化具有保守性(图2).miRNA的合成需要DCL1蛋白和HYL1蛋白的加工(Han et al., 2004; Achkar et al., 2018; Wei et al., 2021b).HYL1-DCL1的互作关系可以追溯到第一批陆地植物中, 来自祖先开花植物的HYL1蛋白可以驱动产生接近原生的miRNA, 以恢复拟南芥HYL1敲除型植株HYL1蛋白的合成(Silvestro et al., 2021; Jia et al., 2021).参与miRNA发生过程的其它家族蛋白在物种间也表现出强烈的保守性(Murphy et al., 2008).研究发现, 高等植物中几乎所有的miRNA生物发生因子也存在于苔藓植物中, 这表明陆地植物进化早期就出现了miRNA (Pietrykowska et al., 2022; Velandia-Huerto et al., 2022).因此, miRNA家族在植物物种进化中持续存在, 是植物基因组中最保守的元素之一.这表明某些miRNA的功能也可能保守, 在多种植物中发挥类似的作用.目前, 已鉴定出许多响应非生物胁迫的miRNA, 涵盖植物应对非生物胁迫的多种途径, 主要包括调节生长发育、营养吸收与分配和信号转导及减少氧化应激损伤(表2; 图3). ... Blocking miR528 function promotes tillering and regrowth in switchgrass 1 2024 ... miR528在植物体内靶向多种抗氧化系统相关酶基因(AO、LAC和SOD等), 参与维持氧化还原稳态以应对非生物胁迫(Liu et al., 2015; Han et al., 2024).SPL9与miR528启动子区域内的特定基序表现出高亲和力, 可激活miR528表达, 调节其介导的L-抗坏血酸氧化酶(AO)降解过程, 形成SPL9-miR528-AO模块, 增强植物对非生物胁迫的抗性(Wu et al., 2017; Yao et al., 2019, 2022).盐胁迫诱导水稻miR528表达, AO丰度下降, 减轻抗坏血酸(AsA)氧化从而清除活性氧积累, 增强水稻对盐胁迫的耐受性(Wang et al., 2021b).低温胁迫使香蕉(Musa acuminata) miR528表达下调, 靶基因PPO转录本丰度剧烈增加, 参与调节植物的抗氧化能力(Zhu et al., 2020). ... Overexpression of Os-microRNA408 enhances drought tolerance in perennial ryegrass 1 2021 ... miR408靶向质体蓝蛋白基因(PCs)以及漆酶基因(LACs)等, 维持电子传递链稳定性, 减少体内活性氧生成, 同时增强超氧化物歧化酶(SOD)和过氧化物酶(POD)等抗氧化酶活性, 以降低活性氧积累水平, 通过提高氧化应激耐受性响应非生物胁迫(Gao et al., 2022; Hu et al., 2023).低温胁迫下拟南芥miR408丰度增高, 靶基因LAC3表达量降低, 诱导抗氧化相关基因表达, 体内活性氧水平降低(Ma et al., 2015).在水稻中过表达miR408下调靶基因质体蓝蛋白基因, 导致离子泄漏率降低, SOD酶活性升高, 增强低温胁迫耐受性, 提高植物存活率(Sun et al., 2018).将丹参(Salvia miltiorrhiza) miR408前体及启动子序列在本氏烟草(N. benthamiana)中异源表达, 发现在盐胁迫下转基因miR408过表达烟草植株抗氧化基因(NbSOD、NbPOD和NbCAT)的转录水平及酶活性提高, 活性氧积累水平降低, 抗氧化能力提升, 耐盐性增强(Guo et al., 2018).砷胁迫下miR408过表达植株砷积累量降低, 重金属耐受性增强(Kumar et al., 2023).对干旱胁迫下多年生黑麦草(Lolium perenne)叶片进行二氨基联苯胺(DAB)染色, 发现miR408过表达植株叶片染色较浅, 说明其活性氧积累水平降低, 受干旱胁迫引起的氧化应激程度较低(Hang et al., 2021). ... Genome-wide identification and characterization of major RNAi genes highlighting their associated factors in cowpea (Vigna unguiculata (L.) Walp 1 2023 ... 植物miR396-GRFs调控模块影响多个重要的生物学过程.双荧光素酶报告基因实验结果表明, ped-miR396d可以与PeGRF6同源物结合(Li et al., 2023a).构建miR396丰度降低和升高型拟南芥转化体, 发现miR396降低型植株鲜重和主根长等表型评估参数都得到增强, 而miR396升高型植株鲜重和主根长等均有所降低, 营养生长严重延迟, 施加盐胁迫处理后, 降低型拟南芥光合色素含量相比野生型和miR396升高型植株更高, 对盐胁迫耐受性更强(Pegler et al., 2021a).缺氮条件下, 水稻miR396敲除型植株地上部生物量相较野生型增加25%, 氮转运和代谢相关基因表达上调, miR396的敲除导致植株氮利用率提高, 与其靶基因OsGRF4过表达植株表型一致, 说明miR396调节植物氮同化及氮代谢(Zhang et al., 2020b), 低磷条件下也有相似的实验结果(Pegler et al., 2021b).此外, 众多研究表明, miR396在火龙果(Hylocereus polyrhizus)、菊花(Chrysanthemum indicum)、凤丹(Paeonia ostii)、番茄、苜蓿、花生(Arachis hypogaea)和豇豆(Vigna unguiculata)等多个物种中均能通过促进水分及营养元素吸收、增加光合色素含量和增加籽粒产量和质量等途径增强植物对非生物胁迫的耐受性(Pegler et al., 2020; Fracasso et al., 2021; Liu et al., 2021; Cai et al., 2022; Guo et al., 2022a; Zhang et al., 2022; Hasan et al., 2023; Ding et al., 2024). ... Research progress on miR396-GRF module regulating plant stress response 2 2022 ... Target genes and functions of miRNAs in plant response to abiotic stresses
非生物胁迫可上调或下调miRNA在植物中的表达水平.在150 mmol·L-1 NaCl胁迫下, 苹果miR156a的表达随胁迫时间的推移而降低(Ma et al., 2021).番茄和拟南芥中的miR169受高温胁迫诱导上调表达(Rao et al., 2022).干旱胁迫显著上调水稻旗叶和根系中miR408及其前体pre-miR408的丰度(Balyan et al., 2023).但相同miRNA在不同物种中的表达也存在差异.干旱胁迫诱导水稻(Lu et al., 2018)和拟南芥(Liu et al., 2008) miR393表达上调, 但miR393在高粱(Sorghum bicolor) (Hamza et al., 2016)和野生桔梗(Ipomoea campanulata) (Ghorecha et al., 2014)中的表达却受到干旱抑制, 大麦(Hordeum vulgare) miR393过表达植株表现为对干旱更加敏感(Yuan et al., 2019b; Jiang et al., 2022a).盐胁迫下, miR396在西伯利亚翦股颖(Agrostis stolonifera) (Yuan et al., 2019a)、番茄和烟草(Nicotiana tabacum) (Chen et al., 2015)中表达量增高, 在水稻(Gao et al., 2010)、旱柳(Salix matsudana) (Zhou et al., 2012)和拟南芥(Pegler et al., 2021a)中表达量降低.在同种非生物胁迫下, 相同miRNA在不同植物物种中表达模式相反, 表明miRNA在不同植物响应相同非生物胁迫时可能发挥不同的作用.未来仍需要对非生物胁迫下植物miRNA进行大规模鉴定及表达模式分析, 从而进一步阐明miRNA调控网络在植物体内的运行机制. ... Fossil data support a pre-cretaceous origin of flowering plants 1 2021 ... 多种植物中含有大多数的miRNA家族, miRNA在物种中的进化具有保守性(图2).miRNA的合成需要DCL1蛋白和HYL1蛋白的加工(Han et al., 2004; Achkar et al., 2018; Wei et al., 2021b).HYL1-DCL1的互作关系可以追溯到第一批陆地植物中, 来自祖先开花植物的HYL1蛋白可以驱动产生接近原生的miRNA, 以恢复拟南芥HYL1敲除型植株HYL1蛋白的合成(Silvestro et al., 2021; Jia et al., 2021).参与miRNA发生过程的其它家族蛋白在物种间也表现出强烈的保守性(Murphy et al., 2008).研究发现, 高等植物中几乎所有的miRNA生物发生因子也存在于苔藓植物中, 这表明陆地植物进化早期就出现了miRNA (Pietrykowska et al., 2022; Velandia-Huerto et al., 2022).因此, miRNA家族在植物物种进化中持续存在, 是植物基因组中最保守的元素之一.这表明某些miRNA的功能也可能保守, 在多种植物中发挥类似的作用.目前, 已鉴定出许多响应非生物胁迫的miRNA, 涵盖植物应对非生物胁迫的多种途径, 主要包括调节生长发育、营养吸收与分配和信号转导及减少氧化应激损伤(表2; 图3). ... Role of abiotic stress responsive miRNAs in Arabidopsis root development 1 2020 ... miR169是植物响应非生物胁迫的普遍调节因子, 通过转录本切割拟制转录因子Nuclear Factor-Y亚基A编码基因家族(NF-YA)的表达(Rao et al., 2020; Wang et al., 2023a), 在植物生长发育中起重要作用.干旱和盐胁迫下, 玉米幼苗miR169与NF-YA基因的表达呈相反趋势, 符合miRNA靶向调控特征(Luan et al., 2015).干旱和盐胁迫可诱导甘蓝型油菜(B. napus)叶片和根系中miR169的丰度变化, 将Bna- miR169和BnaNF-YA在拟南芥中异源表达后, 发现与未经转化的拟南芥相比, 转化Bna-miR169植株在非生物胁迫下表现出根系更短及种子萌发延迟, NF-YA表达水平下调; 而转化BnaNF-YA植株则根系伸长, 种子萌发率更高(Wang et al., 2022).杨树(Populus trichocarpa) miR169过表达植株在正常生长条件下表现为芽和根系伸长, 在干旱胁迫下过表达miR169显著提高杨树的光合速率, 降低失水率, 增强其抗旱性.而NF-YA6过表达植株则表现出生长减缓以及对干旱环境敏感(Jiao et al., 2021).miR169及其靶基因参与调节主根长度和根尖分生组织活性(Sorin et al., 2014; Singh et al., 2020).对NF-YA过表达拟南芥进行功能和转录组分析, 发现其差异表达基因集中在碳代谢和细胞壁的生物合成与修饰(Leyva-Gonzalez et al., 2012). ... 1 2017 ... miR166是植物应对非生物胁迫的重要调节因子, 负调控转录因子HD-Zip III家族表达, 温度、干旱和盐等非生物胁迫均能诱导miR166表达(Zhang et al., 2018; Yadav et al., 2021, 2023; Li et al., 2022; Tian et al., 2022).生长素处理诱导miR166转录本丰度降低, 赤霉素、脱落酸、细胞分裂素、茉莉酸和水杨酸处理诱导miR166强烈表达, 不同激素不敏感型突变体表达模式与施加外源激素相反, 且miR166过表达植株通过对HD-Zip IIIs转录后调控改变植物对激素变化的敏感性, 表明miR166-HD-Zip IIIs调控模块对于维持植物体内激素信号转导平衡具有重要意义(Singh et al., 2017).对野生型和miR166 STTM型植株进行差异表达基因分析, 发现一些激素相关基因显著差异表达, 包括脱落酸诱导蛋白基因、生长素转运蛋白基因、吲哚-3-丙酮酸单加氧酶基因等.miR166 STTM型植株上调脱落酸生物合成及信号转导蛋白基因的表达, 降低生长素转运蛋白编码基因的表达水平, 表现为脱落酸含量增加, 生长素含量降低, 协同调节植物对非生物胁迫的耐受性(Li et al., 2020b). ... Diurnal oscillation in the accumulation of Arabidopsis microRNAs, miR167, miR168, miR171 and miR398 1 2009 ... miR171启动子上存在光、非生物胁迫以及与胁迫相关的外源激素信号响应元件, 在转录后水平调控SCL转录因子表达(Li et al., 2021; Pei et al., 2023).拟南芥miR171家族受盐、温度和干旱等非生物胁迫信号调控, 过表达株系生物量和抗氧化酶活性均高于野生型, 表现出更强的抗性(Liu et al., 2008; Sun et al., 2022; Han and Zhou, 2022).光信号诱导miR171表达, miR171的表达丰度在光照期增高, 黑暗期降低(Sire et al., 2009).生长素诱导miR171通过靶向PyrSCL6和PyrSCL22负调控生长素信号级联, 促进植物生长发育(Jiang et al., 2018); Sly-miR171沉默株系及靶基因SlGRAS24过表达株系中许多赤霉素和生长素相关基因下调, 表明番茄miR171可调节赤霉素以及生长素信号转导(Huang et al., 2017).水稻miR171/SCL6模块调控类黄酮生物合成, 干旱胁迫下过表达植株叶片温度较低, 茎秆高度及谷物产量更高, 抗旱性增强(Um et al., 2022). ... Response of microRNAs to cold treatment in the young spikes of common wheat 1 2017 ... Sequencing and identification of plant abiotic stress response miRNAs
miR156/SPL模块可提高植物对非生物胁迫的耐受性(Wang et al., 2023c; Yuan et al., 2023).利用RLM-5′ RACE技术分析苹果miR156与MdSPL13的关系, 发现MdSPL13的3′UTR区域存在miR156靶标, 证实苹果miR156的靶点为MdSPL13.进一步验证表明, MdSPL13靶向MdWRKY100基因启动子, 因此在苹果中miR156/SPL模块通过靶向调控MdWRKY100调节其耐盐性(Ma et al., 2021).紫花苜蓿(Medicago sativa) miR156过表达植株与野生型相比, 地上部生长更加茂盛, 产量及品质更高, 在盐胁迫下过表达植株中盐含量显著低于野生型, 表现出更强的耐盐性(Wang et al., 2021a).对SPL13 RNAi植株施加干旱胁迫后, 发现叶片中差异表达的基因多集中在光反应系统中, 包括光系统I、II和电子传递链, 说明miR156在叶片中可靶向SPL13调节植物的光合作用, 以抵御干旱胁迫(Feyissa et al., 2020).miR156还靶向SPL2、SPL10、SPL16、SPL18和SPL23等SPL家族基因, 参与植物根系发育、营养生长及生长阶段的转变等重要生命活动(Barrera-Rojas et al., 2020; Su et al., 2023; Zhou and Yarra, 2023; Wang et al., 2023b; Wei et al., 2024). ... The action of ARGONAUTE1 in the miRNA pathway and its regulation by the miRNA pathway are crucial for plant development 1 2004 ... 植物miRNA在细胞核中合成(O'Brien et al., 2018; Bajczyk et al., 2023)(图1).高等植物中大多数miRNA基因代表独立的转录单元, 存在于基因间, 少数存在于蛋白质或非编码基因的内含子中(Millar and Waterhouse, 2005; Stepien et al., 2017; Pietrykowska et al., 2022).迄今为止, 已证实植物miRNA是由miRNA基因在RNA聚合酶II (RNA Pol-II)的作用下转录合成.在该酶的作用下, miRNA基因转录成具有5′端加帽和3′端多聚腺苷酸化的初级转录本(pri-miRNA), pri- miRNA折叠形成发夹状茎环结构(stem-loop).pri- miRNA被DCL1蛋白(DICER-LIKE1)或其它蛋白识别切割为前体miRNA (pre-miRNA), 并在双链RNA结合蛋白HYL1 (Hyponastic Leaves1)和锌指蛋白SE (Serrate)的作用下被进一步加工成miRNA/miRNA*复合体(Park et al., 2002; Lobbes et al., 2006; Dong et al., 2008; Bologna et al., 2009; Wang et al., 2019; Li and Yu, 2021; Xie et al., 2021).随后经甲基转移酶HEN1 (Hua Enhancer1)甲基化修饰3′端后与AGO蛋白结合, 去除miRNA*, 仅留下miRNA稳定在RNA介导的沉默复合物(RNA-induced silencing complex, RISC)中(Vaucheret et al., 2004; Yang et al., 2006; Baranauske et al., 2015; Tomassi et al., 2020; 武亮和戚益军, 2020).但miRNA从细胞核输出到细胞质中的过程仍不完全明确(Iki et al., 2010; Yu et al., 2017; Zhang et al., 2020a).miRNA与AGO蛋白结合后通过CRM1/EXPO1通路输出到细胞质中(Bologna et al., 2018), 然而也有部分研究发现miRNA存在以双链形式输出到细胞质中的可能性(Dalmadi et al., 2019).有研究表明, 与AGO蛋白结合输出到细胞质中的miRNA大多以转录后的方式形成, 而大部分以双链形式输出到细胞质中的miRNA以共转录的方式形成(Gonzalo et al., 2022). ... 1 2022 ... 多种植物中含有大多数的miRNA家族, miRNA在物种中的进化具有保守性(图2).miRNA的合成需要DCL1蛋白和HYL1蛋白的加工(Han et al., 2004; Achkar et al., 2018; Wei et al., 2021b).HYL1-DCL1的互作关系可以追溯到第一批陆地植物中, 来自祖先开花植物的HYL1蛋白可以驱动产生接近原生的miRNA, 以恢复拟南芥HYL1敲除型植株HYL1蛋白的合成(Silvestro et al., 2021; Jia et al., 2021).参与miRNA发生过程的其它家族蛋白在物种间也表现出强烈的保守性(Murphy et al., 2008).研究发现, 高等植物中几乎所有的miRNA生物发生因子也存在于苔藓植物中, 这表明陆地植物进化早期就出现了miRNA (Pietrykowska et al., 2022; Velandia-Huerto et al., 2022).因此, miRNA家族在植物物种进化中持续存在, 是植物基因组中最保守的元素之一.这表明某些miRNA的功能也可能保守, 在多种植物中发挥类似的作用.目前, 已鉴定出许多响应非生物胁迫的miRNA, 涵盖植物应对非生物胁迫的多种途径, 主要包括调节生长发育、营养吸收与分配和信号转导及减少氧化应激损伤(表2; 图3). ... Salinity tolerance in plants. Quantitative approach to ion transport starting from halophytes and stepping to genetic and protein engineering for manipulating ion fluxes 1 2015 ... 植物在其生命周期中可能会面临因外界环境改变导致的各种非生物胁迫(干旱、温度、盐和重金属等), 影响植物体内的各种生命活动, 如活性氧调节(Liu et al., 2022)、激素信号转导(Du et al., 2013)、电解质稳定以及细胞膜完整性(Hou et al., 2016; Muchate et al., 2016; 王凯悦等, 2018), 最终影响植物生长发育进程, 使其产量及品质下降.近年来, 全球气候变化加剧了非生物胁迫发生的频率及危害程度.有研究表明, 全球遭受干旱胁迫的农田比例高达45%, 世界上约38%的人口饱受因盐胁迫导致的作物减产及品质下降困扰(Volkov, 2015; Rhaman et al., 2021).因此, 增强植物对非生物胁迫的耐受性至关重要. ... Rice SPX- major facility Superfamily3, a vacuolar phosphate efflux transporter, is involved in maintaining phosphate homeostasis in rice 1 2015 ... miR827可靶向含SPX结构域的基因, 但不同物种中靶基因的进化存在差异.拟南芥miR827靶向氮限制适应基因(NLA), 水稻miR827靶向磷酸盐转运蛋白基因(PHT5), 调节磷酸盐的运输和储存(Hsieh et al., 2009; Lin et al., 2010, 2018; Wang et al., 2015; Liu et al., 2016).缺磷条件下miR827受到强烈诱导以响应磷酸盐的调控过程(Hackenberg et al., 2013).烟草miR827在缺磷处理下表达上调, 表现出对磷缺乏的正响应, 与拟南芥和水稻等表达模式一致(顾冕等, 2016).研究发现, 低氮处理下拟南芥miR827表达量增加, 靶基因NLA转录本无显著变化, 但蛋白表达丰度随缺氮时间的延长显著下降, 说明miR827通过抑制靶基因NLA翻译调控植物的氮分配过程(刘文文, 2016). ... Brassica napus miR169 regulates BnaNF-YA in salinity, drought and ABA responses 2 2022 ... Target genes and functions of miRNAs in plant response to abiotic stresses
非生物胁迫可上调或下调miRNA在植物中的表达水平.在150 mmol·L-1 NaCl胁迫下, 苹果miR156a的表达随胁迫时间的推移而降低(Ma et al., 2021).番茄和拟南芥中的miR169受高温胁迫诱导上调表达(Rao et al., 2022).干旱胁迫显著上调水稻旗叶和根系中miR408及其前体pre-miR408的丰度(Balyan et al., 2023).但相同miRNA在不同物种中的表达也存在差异.干旱胁迫诱导水稻(Lu et al., 2018)和拟南芥(Liu et al., 2008) miR393表达上调, 但miR393在高粱(Sorghum bicolor) (Hamza et al., 2016)和野生桔梗(Ipomoea campanulata) (Ghorecha et al., 2014)中的表达却受到干旱抑制, 大麦(Hordeum vulgare) miR393过表达植株表现为对干旱更加敏感(Yuan et al., 2019b; Jiang et al., 2022a).盐胁迫下, miR396在西伯利亚翦股颖(Agrostis stolonifera) (Yuan et al., 2019a)、番茄和烟草(Nicotiana tabacum) (Chen et al., 2015)中表达量增高, 在水稻(Gao et al., 2010)、旱柳(Salix matsudana) (Zhou et al., 2012)和拟南芥(Pegler et al., 2021a)中表达量降低.在同种非生物胁迫下, 相同miRNA在不同植物物种中表达模式相反, 表明miRNA在不同植物响应相同非生物胁迫时可能发挥不同的作用.未来仍需要对非生物胁迫下植物miRNA进行大规模鉴定及表达模式分析, 从而进一步阐明miRNA调控网络在植物体内的运行机制. ... 1 2021a ... miR397可通过碱基互补配对直接切割LACs转录本, 植物LACs是木质素聚合的正调节剂, 能够将单木质醇聚合为木质素, 在细胞壁木质素生物合成和植物抵抗外界不良刺激中发挥重要作用(Bao et al., 1993; Ranocha et al., 2002; Li et al., 2020a; Wei et al., 2021a).镉胁迫下拟南芥miR397过表达植株通过调控LAC2、LAC4和LAC17的表达改变木质素含量, 从而降低植物对镉的耐受性, 而利用短串联靶标模拟(short tandem target mimic, STTM)技术培育的miR397沉默植株木质素含量更高, 细胞壁镉含量和细胞总镉含量更低, 表现出更强的镉耐受性(Ali et al., 2023).在盐胁迫下也有同样的结果, 过表达miR397拟南芥植株对外界盐度变化更敏感(Nguyen et al., 2020).miR397-LAC4/17模块可通过细胞壁木质化调节拟南芥和柳橙中的硼流动, 拟南芥和柳橙LAC4过表达植株分别触发维管系统和木质部细胞壁高硼依赖性木质化, 减少植物体内游离硼含量, 从而提高对硼胁迫的耐受性(Huang et al., 2022). ... Structural basis of microRNA processing by dicer-like 1 1 2021b ... 多种植物中含有大多数的miRNA家族, miRNA在物种中的进化具有保守性(图2).miRNA的合成需要DCL1蛋白和HYL1蛋白的加工(Han et al., 2004; Achkar et al., 2018; Wei et al., 2021b).HYL1-DCL1的互作关系可以追溯到第一批陆地植物中, 来自祖先开花植物的HYL1蛋白可以驱动产生接近原生的miRNA, 以恢复拟南芥HYL1敲除型植株HYL1蛋白的合成(Silvestro et al., 2021; Jia et al., 2021).参与miRNA发生过程的其它家族蛋白在物种间也表现出强烈的保守性(Murphy et al., 2008).研究发现, 高等植物中几乎所有的miRNA生物发生因子也存在于苔藓植物中, 这表明陆地植物进化早期就出现了miRNA (Pietrykowska et al., 2022; Velandia-Huerto et al., 2022).因此, miRNA家族在植物物种进化中持续存在, 是植物基因组中最保守的元素之一.这表明某些miRNA的功能也可能保守, 在多种植物中发挥类似的作用.目前, 已鉴定出许多响应非生物胁迫的miRNA, 涵盖植物应对非生物胁迫的多种途径, 主要包括调节生长发育、营养吸收与分配和信号转导及减少氧化应激损伤(表2; 图3). ... Identification of miR390-TAS3-ARF pathway in response to salt stress in Helianthus tuberosus L 1 2020 ... miR390和TAS3衍生的反式作用短干扰RNA (tasiRNA)和生长素反应因子ARF形成生长素反应调节网络以响应非生物胁迫(Marin et al., 2010; de Felippes et al., 2017; Yu et al., 2020).低温胁迫诱导马铃薯(S. tuberosum) miR390表达上调, 盐胁迫处理使miR390表达下调(谢洁等, 2019).miR390/ TAS3/ARF模块调节不同外界环境中植物对生长素的敏感性, 盐胁迫下过表达miR390植株耐盐性增强, ARF表达丰度降低, 生长素通路中典型基因转录较多, 以维持生长素信号转导(He et al., 2018; Dastidar et al., 2019; Wen et al., 2020; Shi et al., 2023).分析野草莓(Fragaria vesca) miR390启动子序列, 发现其包含生长素反应元件(AuxRE), 施加外源生长素可显著提高miR390的表达水平, 通过miR390-tasiRNA3- ARF4通路参与调控植物的生长发育进程(Dong et al., 2022).研究发现, 棉花(Gossypium hirsutum) miR390也可以靶向切割GhCEPR2, 负调控耐盐性, CEPR2通过磷酸化脱落酸信号转导途径中的蛋白质调节脱落酸含量, 介导植物对盐胁迫的响应(Chu et al., 2022).铜胁迫下, 拟南芥miR390通过反式作用siRNAs的生成调控生长素响应因子, 从而增强植物对重金属的抗性(谢洁等, 2018). ... 1 1993 ... miRNA是一类长约20-24个核苷酸的内源性非编码小分子RNA, 通过剪切靶mRNA和抑制靶基因的翻译, 在转录后水平负调控真核生物基因的表达.miRNA于1993年从线虫(Caenorhabditis elegans)中被发现(Lee et al., 1993; Wightman et al., 1993; Fire et al., 1998), 随后于2002年在模式植物拟南芥(Arabidopsis thaliana)中被鉴定发现(Reinhart et al., 2002).随着高通量测序技术和生物学分析手段的发展, 研究人员陆续从其它物种中鉴定出多个miRNA.目前miRBase (http://mirbase.org/)以及PmiREN2.0 (https://pmiren.com/)等miRNA数据库中涵盖了近300个物种的miRNA序列信息(Kozomara et al., 2019; Guo et al., 2022b). ... miR393 contributes to the embryogenic transition induced in vitro in Arabidopsis via the modification of the tissue sensitivity to auxin treatment 1 2016 ... miR393靶向F-box生长素受体TIR1和生长素信号转导蛋白AFBs基因, 通过释放生长素反应因子(ARF)和介导Aux/IAA阻遏蛋白水解调节生长素信号转导以响应非生物胁迫(Chen et al., 2011; Wojcik and Gaj, 2016; Cai et al., 2017).冷胁迫处理后植物冷响应基因表达增强, 过表达miR393植株相关基因表达量增幅更高, 对低温耐受性更强, 外源生长素处理负调控上述冷响应基因, 说明生长素信号通路参与植物耐寒性, 这表明miR393抑制的生长素信号转导能增强植物的抗寒性(Liu et al., 2017).对铝胁迫下的大麦施加外源生长素可增强Al3+诱导的根系抑制作用, 且Al3+诱导根尖生长素应答基因表达, miR393的过表达可减弱这种抑制作用并下调相关反应基因的表达, 表明miR393通过改变大麦生长素信号输出调控根系对Al的敏感性(Bai et al., 2017).研究发现, miR393调节生长素反应因子进而调控气孔发育相关基因的表达, 过表达植株表现出叶片保卫细胞长度变短, 气孔密度增加, 植株对干旱胁迫更敏感, 而敲除系植株则呈现相反的表型(Yuan et al., 2019b).在匍匐翦股颖和大豆(Glycine max)等植物中也获得了相似的结果(Zhao et al., 2019; Xing et al., 2023).也有研究表明, miR393通过盐胁迫诱导的应激激素脱落酸与生长素之间的拮抗作用参与植物对盐胁迫的响应(Denver and Ullah, 2019). ... ROS accumulation and antiviral defence control by microRNA528 in rice 1 2017 ... miR528在植物体内靶向多种抗氧化系统相关酶基因(AO、LAC和SOD等), 参与维持氧化还原稳态以应对非生物胁迫(Liu et al., 2015; Han et al., 2024).SPL9与miR528启动子区域内的特定基序表现出高亲和力, 可激活miR528表达, 调节其介导的L-抗坏血酸氧化酶(AO)降解过程, 形成SPL9-miR528-AO模块, 增强植物对非生物胁迫的抗性(Wu et al., 2017; Yao et al., 2019, 2022).盐胁迫诱导水稻miR528表达, AO丰度下降, 减轻抗坏血酸(AsA)氧化从而清除活性氧积累, 增强水稻对盐胁迫的耐受性(Wang et al., 2021b).低温胁迫使香蕉(Musa acuminata) miR528表达下调, 靶基因PPO转录本丰度剧烈增加, 参与调节植物的抗氧化能力(Zhu et al., 2020). ... Small RNA, no small feat: plants deploy 22 nt siRNAs to cope with environmental stress 1 2020 ... 植物miRNA在细胞核中合成(O'Brien et al., 2018; Bajczyk et al., 2023)(图1).高等植物中大多数miRNA基因代表独立的转录单元, 存在于基因间, 少数存在于蛋白质或非编码基因的内含子中(Millar and Waterhouse, 2005; Stepien et al., 2017; Pietrykowska et al., 2022).迄今为止, 已证实植物miRNA是由miRNA基因在RNA聚合酶II (RNA Pol-II)的作用下转录合成.在该酶的作用下, miRNA基因转录成具有5′端加帽和3′端多聚腺苷酸化的初级转录本(pri-miRNA), pri- miRNA折叠形成发夹状茎环结构(stem-loop).pri- miRNA被DCL1蛋白(DICER-LIKE1)或其它蛋白识别切割为前体miRNA (pre-miRNA), 并在双链RNA结合蛋白HYL1 (Hyponastic Leaves1)和锌指蛋白SE (Serrate)的作用下被进一步加工成miRNA/miRNA*复合体(Park et al., 2002; Lobbes et al., 2006; Dong et al., 2008; Bologna et al., 2009; Wang et al., 2019; Li and Yu, 2021; Xie et al., 2021).随后经甲基转移酶HEN1 (Hua Enhancer1)甲基化修饰3′端后与AGO蛋白结合, 去除miRNA*, 仅留下miRNA稳定在RNA介导的沉默复合物(RNA-induced silencing complex, RISC)中(Vaucheret et al., 2004; Yang et al., 2006; Baranauske et al., 2015; Tomassi et al., 2020; 武亮和戚益军, 2020).但miRNA从细胞核输出到细胞质中的过程仍不完全明确(Iki et al., 2010; Yu et al., 2017; Zhang et al., 2020a).miRNA与AGO蛋白结合后通过CRM1/EXPO1通路输出到细胞质中(Bologna et al., 2018), 然而也有部分研究发现miRNA存在以双链形式输出到细胞质中的可能性(Dalmadi et al., 2019).有研究表明, 与AGO蛋白结合输出到细胞质中的miRNA大多以转录后的方式形成, 而大部分以双链形式输出到细胞质中的miRNA以共转录的方式形成(Gonzalo et al., 2022). ... 小RNA, 大本领: 22 nt siRNAs在植物适应逆境中的重要作用 1 2020 ... 植物miRNA在细胞核中合成(O'Brien et al., 2018; Bajczyk et al., 2023)(图1).高等植物中大多数miRNA基因代表独立的转录单元, 存在于基因间, 少数存在于蛋白质或非编码基因的内含子中(Millar and Waterhouse, 2005; Stepien et al., 2017; Pietrykowska et al., 2022).迄今为止, 已证实植物miRNA是由miRNA基因在RNA聚合酶II (RNA Pol-II)的作用下转录合成.在该酶的作用下, miRNA基因转录成具有5′端加帽和3′端多聚腺苷酸化的初级转录本(pri-miRNA), pri- miRNA折叠形成发夹状茎环结构(stem-loop).pri- miRNA被DCL1蛋白(DICER-LIKE1)或其它蛋白识别切割为前体miRNA (pre-miRNA), 并在双链RNA结合蛋白HYL1 (Hyponastic Leaves1)和锌指蛋白SE (Serrate)的作用下被进一步加工成miRNA/miRNA*复合体(Park et al., 2002; Lobbes et al., 2006; Dong et al., 2008; Bologna et al., 2009; Wang et al., 2019; Li and Yu, 2021; Xie et al., 2021).随后经甲基转移酶HEN1 (Hua Enhancer1)甲基化修饰3′端后与AGO蛋白结合, 去除miRNA*, 仅留下miRNA稳定在RNA介导的沉默复合物(RNA-induced silencing complex, RISC)中(Vaucheret et al., 2004; Yang et al., 2006; Baranauske et al., 2015; Tomassi et al., 2020; 武亮和戚益军, 2020).但miRNA从细胞核输出到细胞质中的过程仍不完全明确(Iki et al., 2010; Yu et al., 2017; Zhang et al., 2020a).miRNA与AGO蛋白结合后通过CRM1/EXPO1通路输出到细胞质中(Bologna et al., 2018), 然而也有部分研究发现miRNA存在以双链形式输出到细胞质中的可能性(Dalmadi et al., 2019).有研究表明, 与AGO蛋白结合输出到细胞质中的miRNA大多以转录后的方式形成, 而大部分以双链形式输出到细胞质中的miRNA以共转录的方式形成(Gonzalo et al., 2022). ... Progress in the regulation of plant miR397 and their target genes 1 2022 ... Target genes and functions of miRNAs in plant response to abiotic stresses
本文的其它图/表
|

 首页
首页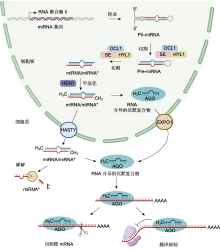
{kind=link}