植物miRNA响应非生物胁迫研究进展
周文杰, 张文瀚, 贾玮, 许自成, 黄五星
植物学报
2024, 59 ( 5):
810-833.
DOI: 10.11983/CBB24020
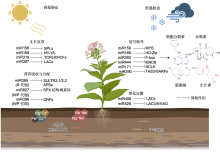
干旱、极端温度、盐和重金属等非生物胁迫导致植物产量和品质下降。miRNA是一类长约20-24个核苷酸的内源性非编码小分子RNA, 通过形成miRNA介导的沉默复合物(RISC)剪切靶mRNA并抑制靶基因的翻译, 在转录后水平负调控真核生物基因表达。高通量测序技术的快速发展使得植物物种中大量响应非生物胁迫的miRNA得到鉴定和表征。非生物胁迫下植物miRNA与其靶基因结合, 构成了控制各种生命活动的大型基因调控网络, 包括生长发育、营养吸收与分配、信号转导与氧化应激, 从而提高植物的抗逆性。深入理解miRNA的功能及其调控机制对于通过基因工程进行作物改良和抗逆育种至关重要。该文综述了近年来miRNA的生物合成及其作用机制研究进展, 重点探讨了参与调控植物响应非生物胁迫miRNA的鉴定及功能, 并展望了该领域可能的研究方向。
| 名称 | 取样部位 | 胁迫处理类型和时间 | miRNA差异
表达数量 | 参考文献 | 拟南芥
(Arabidopsis thaliana) | 全幼苗 | 200 mmol·L-1甘露醇7天(干旱胁迫) | 123 | Pegler et al., 2019 | | 全幼苗 | 昼夜温度为32°C/28°C 7天(高温胁迫) | 121 | Pegler et al., 2019 | | 全幼苗 | 150 mmol·L-1氯化钠7天(盐胁迫) | 118 | Pegler et al., 2019 | | 叶片通气组织 | 放置4°C下4小时(低温胁迫) | 93 | Tiwari et al., 2020 | 水稻
(Oryza sativa) | 叶片组织 | 放置4°C下5小时(低温胁迫) | 23 | Shen et al., 2010 | | 叶片 | 不浇水直至出现典型症状(干旱胁迫) | 64 | Cheah et al., 2015 | | 茎 | 不浇水直至出现典型症状(干旱胁迫) | 71 | Cheah et al., 2015 | | 全幼苗 | 200 mmol·L-1氯化钠2天(盐胁迫) | 164 | Chen et al., 2023 | | 具有乳熟期颖果的小穗 | 昼夜温度为38°C/22°C 6小时(高温胁迫) | 96 | Payne et al., 2023 | | 全幼苗 | 昼夜温度为10°C/8°C 5天(低温胁迫) | 26 | Zhao et al., 2022b | 玉米
(Zea mays) | 每株第4片叶 | 昼夜温度为25°C/4°C, 直至该叶片完全成熟(低温胁迫) | 65 | Aydinoglu, 2020 | | 每株第10片叶中部 | 昼夜温度为40°C/25°C 5天(高温胁迫) | 102 | Zhang et al., 2019 | | 叶片 | 16%聚乙二醇24小时(干旱胁迫) | 68 | Wei et al., 2009 | | 根系 | 200 mmol·L-1氯化钠24小时(盐胁迫) | 98 | Ding et al., 2009 | 马铃薯
(Solanum
tuberosum) | 第3片完全展开的叶片 | 放置0°C下4小时(低温胁迫) | 136 | Yan et al., 2021 | | 全植株 | 去除培养液6小时(干旱胁迫) | 230 | Shin et al., 2017 | | 叶片组织 | 放置35°C下1小时(高温胁迫) | 204 | 张国栋, 2021 | | 根系 | 50 mmol·L-1 NaHCO3 24小时(盐胁迫) | 169 | 康益晨, 2021 | 番茄(S.
lycopersicum) | 长度为6.0-6.5 mm的花蕾雄蕊 | 昼夜温度为35°C/30°C 2天(高温胁迫) | 69 | Pan et al., 2017 | | 长度为6.0-6.5 mm的花蕾雌蕊 | 昼夜温度为35°C/30°C 12天(高温胁迫) | 30 | Pan et al., 2017 | | 根系 | 5%聚乙二醇7天(干旱胁迫) | 101 | Candar-Cakir et al., 2016 | | 茎顶端第3片叶 | 放置4°C下48小时(低温胁迫) | 49 | Cao et al., 2014 | | 根系 | 200 mmol·L-1氯化钠12小时(盐胁迫) | 145 | Wang et al., 2021d | 烟草
(Nicotiana
tabacum) | 叶片 | 放置6°C下24小时(低温胁迫) | 25 | Hu et al., 2019 | | 叶片 | 不浇水10天(干旱胁迫) | 32 | Chen et al., 2017 | | 叶片 | 100 mmol·L-1氯化钠24小时(盐胁迫) | 33 | Xu et al., 2019 | 小麦
(Triticum
aestivum) | 幼穗 | 放置0°C下48小时(低温胁迫) | 39 | Song et al., 2017 | | 叶片组织 | 放置37°C下5天(高温胁迫) | 79 | Ragupathy et al., 2016 | | 旗叶 | 保持土壤含水率为田间容量的一半(6%)
(干旱胁迫) | 54 | Liu et al., 2020 | | 全植株 | 150 mmol·L-1氯化钠24小时(盐胁迫) | 222 | Feng et al., 2017 | | 根和叶组织 | 脱水处理8小时(干旱胁迫) | 141 | Kantar et al., 2011 | | 冠状组织 | 自然条件下冬季生育期(低温胁迫) | 92 | Wang et al., 2021c |
View table in article
表1
响应植物非生物胁迫miRNAs的测序鉴定
正文中引用本图/表的段落
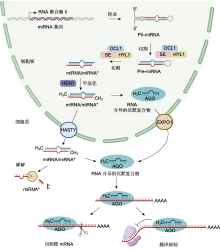
随着微阵列和高通量深度测序的快速发展, 研究人员在植物响应非生物胁迫方面鉴定出大量miRNA。对不同植物进行干旱、温度和盐等胁迫处理后鉴定响应相关胁迫的miRNA (表1)。研究发现, 对不同品种番茄(Solanum lycopersicum)幼苗进行干旱胁迫处理后, 分别以根系和地上部为材料建立小RNA文库进行深度测序, 鉴定出来自578个家族的699个miRNAs, 其中干旱耐受型品种在根系中差异表达的miRNA为101个, 在地上部中差异表达的miRNA为58个, 干旱敏感型品种在根系和地上部中差异表达的miRNA分别为46和61个(Candar-Cakir et al., 2016)。拟南芥幼苗在干旱、高温和盐胁迫处理下相较对照组差异表达的miRNA分别为123、121和118个, 包含特异性响应干旱胁迫miRNA 33个、特异性响应盐胁迫miRNA 25个、特异性响应高温胁迫miRNA 24个以及在三者中共同响应的miRNA 52个(Pegler et al., 2019)。这表明非生物胁迫下植物miRNA的鉴定结果受物种、品种、胁迫处理方法、处理时间及组织特异性等因素影响。
非生物胁迫可上调或下调miRNA在植物中的表达水平.在150 mmol·L-1 NaCl胁迫下, 苹果miR156a的表达随胁迫时间的推移而降低(Ma et al., 2021).番茄和拟南芥中的miR169受高温胁迫诱导上调表达(Rao et al., 2022).干旱胁迫显著上调水稻旗叶和根系中miR408及其前体pre-miR408的丰度(Balyan et al., 2023).但相同miRNA在不同物种中的表达也存在差异.干旱胁迫诱导水稻(Lu et al., 2018)和拟南芥(Liu et al., 2008) miR393表达上调, 但miR393在高粱(Sorghum bicolor) (Hamza et al., 2016)和野生桔梗(Ipomoea campanulata) (Ghorecha et al., 2014)中的表达却受到干旱抑制, 大麦(Hordeum vulgare) miR393过表达植株表现为对干旱更加敏感(Yuan et al., 2019b; Jiang et al., 2022a).盐胁迫下, miR396在西伯利亚翦股颖(Agrostis stolonifera) (Yuan et al., 2019a)、番茄和烟草(Nicotiana tabacum) (Chen et al., 2015)中表达量增高, 在水稻(Gao et al., 2010)、旱柳(Salix matsudana) (Zhou et al., 2012)和拟南芥(Pegler et al., 2021a)中表达量降低.在同种非生物胁迫下, 相同miRNA在不同植物物种中表达模式相反, 表明miRNA在不同植物响应相同非生物胁迫时可能发挥不同的作用.未来仍需要对非生物胁迫下植物miRNA进行大规模鉴定及表达模式分析, 从而进一步阐明miRNA调控网络在植物体内的运行机制. ... MiR393-mediated auxin signaling regulation is involved in root elongation inhibition in response to toxic aluminum stress in barley 1 2017 ... miR393靶向F-box生长素受体TIR1和生长素信号转导蛋白AFBs基因, 通过释放生长素反应因子(ARF)和介导Aux/IAA阻遏蛋白水解调节生长素信号转导以响应非生物胁迫(Chen et al., 2011; Wojcik and Gaj, 2016; Cai et al., 2017).冷胁迫处理后植物冷响应基因表达增强, 过表达miR393植株相关基因表达量增幅更高, 对低温耐受性更强, 外源生长素处理负调控上述冷响应基因, 说明生长素信号通路参与植物耐寒性, 这表明miR393抑制的生长素信号转导能增强植物的抗寒性(Liu et al., 2017).对铝胁迫下的大麦施加外源生长素可增强Al3+诱导的根系抑制作用, 且Al3+诱导根尖生长素应答基因表达, miR393的过表达可减弱这种抑制作用并下调相关反应基因的表达, 表明miR393通过改变大麦生长素信号输出调控根系对Al的敏感性(Bai et al., 2017).研究发现, miR393调节生长素反应因子进而调控气孔发育相关基因的表达, 过表达植株表现出叶片保卫细胞长度变短, 气孔密度增加, 植株对干旱胁迫更敏感, 而敲除系植株则呈现相反的表型(Yuan et al., 2019b).在匍匐翦股颖和大豆(Glycine max)等植物中也获得了相似的结果(Zhao et al., 2019; Xing et al., 2023).也有研究表明, miR393通过盐胁迫诱导的应激激素脱落酸与生长素之间的拮抗作用参与植物对盐胁迫的响应(Denver and Ullah, 2019). ... Recent insights into plant miRNA biogenesis: multiple layers of miRNA level regulation 1 2023 ... 植物miRNA在细胞核中合成(O'Brien et al., 2018; Bajczyk et al., 2023)(图1).高等植物中大多数miRNA基因代表独立的转录单元, 存在于基因间, 少数存在于蛋白质或非编码基因的内含子中(Millar and Waterhouse, 2005; Stepien et al., 2017; Pietrykowska et al., 2022).迄今为止, 已证实植物miRNA是由miRNA基因在RNA聚合酶II (RNA Pol-II)的作用下转录合成.在该酶的作用下, miRNA基因转录成具有5′端加帽和3′端多聚腺苷酸化的初级转录本(pri-miRNA), pri- miRNA折叠形成发夹状茎环结构(stem-loop).pri- miRNA被DCL1蛋白(DICER-LIKE1)或其它蛋白识别切割为前体miRNA (pre-miRNA), 并在双链RNA结合蛋白HYL1 (Hyponastic Leaves1)和锌指蛋白SE (Serrate)的作用下被进一步加工成miRNA/miRNA*复合体(Park et al., 2002; Lobbes et al., 2006; Dong et al., 2008; Bologna et al., 2009; Wang et al., 2019; Li and Yu, 2021; Xie et al., 2021).随后经甲基转移酶HEN1 (Hua Enhancer1)甲基化修饰3′端后与AGO蛋白结合, 去除miRNA*, 仅留下miRNA稳定在RNA介导的沉默复合物(RNA-induced silencing complex, RISC)中(Vaucheret et al., 2004; Yang et al., 2006; Baranauske et al., 2015; Tomassi et al., 2020; 武亮和戚益军, 2020).但miRNA从细胞核输出到细胞质中的过程仍不完全明确(Iki et al., 2010; Yu et al., 2017; Zhang et al., 2020a).miRNA与AGO蛋白结合后通过CRM1/EXPO1通路输出到细胞质中(Bologna et al., 2018), 然而也有部分研究发现miRNA存在以双链形式输出到细胞质中的可能性(Dalmadi et al., 2019).有研究表明, 与AGO蛋白结合输出到细胞质中的miRNA大多以转录后的方式形成, 而大部分以双链形式输出到细胞质中的miRNA以共转录的方式形成(Gonzalo et al., 2022). ... Delineating the tissue-mediated drought stress governed tuning of conserved miR408 and its targets in rice 1 2023 ... 非生物胁迫可上调或下调miRNA在植物中的表达水平.在150 mmol·L-1 NaCl胁迫下, 苹果miR156a的表达随胁迫时间的推移而降低(Ma et al., 2021).番茄和拟南芥中的miR169受高温胁迫诱导上调表达(Rao et al., 2022).干旱胁迫显著上调水稻旗叶和根系中miR408及其前体pre-miR408的丰度(Balyan et al., 2023).但相同miRNA在不同物种中的表达也存在差异.干旱胁迫诱导水稻(Lu et al., 2018)和拟南芥(Liu et al., 2008) miR393表达上调, 但miR393在高粱(Sorghum bicolor) (Hamza et al., 2016)和野生桔梗(Ipomoea campanulata) (Ghorecha et al., 2014)中的表达却受到干旱抑制, 大麦(Hordeum vulgare) miR393过表达植株表现为对干旱更加敏感(Yuan et al., 2019b; Jiang et al., 2022a).盐胁迫下, miR396在西伯利亚翦股颖(Agrostis stolonifera) (Yuan et al., 2019a)、番茄和烟草(Nicotiana tabacum) (Chen et al., 2015)中表达量增高, 在水稻(Gao et al., 2010)、旱柳(Salix matsudana) (Zhou et al., 2012)和拟南芥(Pegler et al., 2021a)中表达量降低.在同种非生物胁迫下, 相同miRNA在不同植物物种中表达模式相反, 表明miRNA在不同植物响应相同非生物胁迫时可能发挥不同的作用.未来仍需要对非生物胁迫下植物miRNA进行大规模鉴定及表达模式分析, 从而进一步阐明miRNA调控网络在植物体内的运行机制. ... A laccase associated with lignification in loblolly pine xylem 1 1993 ... miR397可通过碱基互补配对直接切割LACs转录本, 植物LACs是木质素聚合的正调节剂, 能够将单木质醇聚合为木质素, 在细胞壁木质素生物合成和植物抵抗外界不良刺激中发挥重要作用(Bao et al., 1993; Ranocha et al., 2002; Li et al., 2020a; Wei et al., 2021a).镉胁迫下拟南芥miR397过表达植株通过调控LAC2、LAC4和LAC17的表达改变木质素含量, 从而降低植物对镉的耐受性, 而利用短串联靶标模拟(short tandem target mimic, STTM)技术培育的miR397沉默植株木质素含量更高, 细胞壁镉含量和细胞总镉含量更低, 表现出更强的镉耐受性(Ali et al., 2023).在盐胁迫下也有同样的结果, 过表达miR397拟南芥植株对外界盐度变化更敏感(Nguyen et al., 2020).miR397-LAC4/17模块可通过细胞壁木质化调节拟南芥和柳橙中的硼流动, 拟南芥和柳橙LAC4过表达植株分别触发维管系统和木质部细胞壁高硼依赖性木质化, 减少植物体内游离硼含量, 从而提高对硼胁迫的耐受性(Huang et al., 2022). ... Functional mapping of the plant small RNA methyltransferase: HEN1 physically interacts with HYL1 and DICER-LIKE 1 proteins 1 2015 ... 植物miRNA在细胞核中合成(O'Brien et al., 2018; Bajczyk et al., 2023)(图1).高等植物中大多数miRNA基因代表独立的转录单元, 存在于基因间, 少数存在于蛋白质或非编码基因的内含子中(Millar and Waterhouse, 2005; Stepien et al., 2017; Pietrykowska et al., 2022).迄今为止, 已证实植物miRNA是由miRNA基因在RNA聚合酶II (RNA Pol-II)的作用下转录合成.在该酶的作用下, miRNA基因转录成具有5′端加帽和3′端多聚腺苷酸化的初级转录本(pri-miRNA), pri- miRNA折叠形成发夹状茎环结构(stem-loop).pri- miRNA被DCL1蛋白(DICER-LIKE1)或其它蛋白识别切割为前体miRNA (pre-miRNA), 并在双链RNA结合蛋白HYL1 (Hyponastic Leaves1)和锌指蛋白SE (Serrate)的作用下被进一步加工成miRNA/miRNA*复合体(Park et al., 2002; Lobbes et al., 2006; Dong et al., 2008; Bologna et al., 2009; Wang et al., 2019; Li and Yu, 2021; Xie et al., 2021).随后经甲基转移酶HEN1 (Hua Enhancer1)甲基化修饰3′端后与AGO蛋白结合, 去除miRNA*, 仅留下miRNA稳定在RNA介导的沉默复合物(RNA-induced silencing complex, RISC)中(Vaucheret et al., 2004; Yang et al., 2006; Baranauske et al., 2015; Tomassi et al., 2020; 武亮和戚益军, 2020).但miRNA从细胞核输出到细胞质中的过程仍不完全明确(Iki et al., 2010; Yu et al., 2017; Zhang et al., 2020a).miRNA与AGO蛋白结合后通过CRM1/EXPO1通路输出到细胞质中(Bologna et al., 2018), 然而也有部分研究发现miRNA存在以双链形式输出到细胞质中的可能性(Dalmadi et al., 2019).有研究表明, 与AGO蛋白结合输出到细胞质中的miRNA大多以转录后的方式形成, 而大部分以双链形式输出到细胞质中的miRNA以共转录的方式形成(Gonzalo et al., 2022). ... MiR156-targeted SPL10 controls Arabidopsis root meristem activity and root-derived de novo shoot regeneration via cytokinin responses 1 2020 ... miR156/SPL模块可提高植物对非生物胁迫的耐受性(Wang et al., 2023c; Yuan et al., 2023).利用RLM-5′ RACE技术分析苹果miR156与MdSPL13的关系, 发现MdSPL13的3′UTR区域存在miR156靶标, 证实苹果miR156的靶点为MdSPL13.进一步验证表明, MdSPL13靶向MdWRKY100基因启动子, 因此在苹果中miR156/SPL模块通过靶向调控MdWRKY100调节其耐盐性(Ma et al., 2021).紫花苜蓿(Medicago sativa) miR156过表达植株与野生型相比, 地上部生长更加茂盛, 产量及品质更高, 在盐胁迫下过表达植株中盐含量显著低于野生型, 表现出更强的耐盐性(Wang et al., 2021a).对SPL13 RNAi植株施加干旱胁迫后, 发现叶片中差异表达的基因多集中在光反应系统中, 包括光系统I、II和电子传递链, 说明miR156在叶片中可靶向SPL13调节植物的光合作用, 以抵御干旱胁迫(Feyissa et al., 2020).miR156还靶向SPL2、SPL10、SPL16、SPL18和SPL23等SPL家族基因, 参与植物根系发育、营养生长及生长阶段的转变等重要生命活动(Barrera-Rojas et al., 2020; Su et al., 2023; Zhou and Yarra, 2023; Wang et al., 2023b; Wei et al., 2024). ... 1 2022 ... miR319可调节植物次级细胞壁的形成与发育, 控制叶片及根系生长发育, 从而提高植物对非生物胁迫的耐受性.miR319在植物中靶向调控转录因子TCP4 (Zhang et al., 2023), TCP4可结合VND7启动子并激活其表达, 从而触发与次级细胞壁生物合成和程序性细胞死亡相关的VND7转录网络, 发挥开启植物次级细胞壁合成的开关作用(Sun et al., 2017).拟南芥miR319缺失突变体中TCP4表达量增加, 根尖分生组织被破坏, 根系细胞增殖减少, 导致植株根系伸长减慢, 根系较短(Baulies et al., 2022).水稻miR319过表达植株诱导至少4个靶基因(AsPCF5、AsPCF6、AsPCF8和AsTCP14)表达量降低, 在干旱和盐胁迫下表现出更强的耐受性(Zhou et al., 2013; Zhou and Luo, 2014).在低温胁迫下, miR319在番茄(Valiollahi et al., 2014)、甘蔗(Saccharum officinarum) (Thiebaut et al., 2012)和芥菜(Brassica juncea) (Joshi et al., 2021)中上调表达, 通过调控植物发育应对胁迫. ... Arabidopsis ARGONAUT1 is an RNA slicer that selectively recruits microRNAs and short interfering RNAs 1 2005 ... miRNA主要通过介导靶mRNA剪切和抑制靶基因翻译两种方式在转录后水平发挥作用(Yu et al., 2017; 张翠桔等, 2020).在植物中, miRNA与其对应的靶mRNA序列互补程度较高, 因此植物miRNA发挥作用的主要方式是对靶mRNA进行剪切(Iwakawa and Tomari, 2013).模式植物拟南芥大部分miRNA可以剪切其靶mRNA (German et al., 2008).AGO蛋白具有核酸内切酶活性, 可以与miRNA组装成miRNA- RISC复合体, 识别并结合与其互补配对的靶mRNA并在特定位点特异性切割核苷酸之间的磷酸二酯键(Baumberger and Baulcombe, 2005; Qi et al., 2005; Höeck and Meister, 2008; Mi et al., 2008).mRNA经剪切后断裂成含帽子结构的5′端和polyA尾的3′端切割产物, 进入核酸外切酶降解途径(Elbashir et al., 2001; Llave et al., 2002; Souret et al., 2004; Ren et al., 2014). ... MRNA degradation by miRNAs and GW182 requires both CCR4: NOT deadenylase and DCP1: DCP2 decapping complexes 1 2006 ... miRNA的另一种作用途径是抑制靶基因的翻译过程, 此途径在动物中较为常见, 在植物中的研究相对较少.动物miRNA仅能在种子序列区域(seed region)与靶基因3′端非编码区进行配对, 互补程度不高, 无法直接剪切mRNA, 因此抑制靶基因翻译是动物miRNA的常见作用模式(Huntzinger and Izaurralde, 2011).其作用机制为动物miRNA-RISC复合体除AGO蛋白外, 还含有GW182蛋白, 后者可以与翻译过程中众多因子相互作用.GW182蛋白可以与多腺苷酸结合蛋白相互作用, 使polyA在mRNA 3′端解离, 也可以与相关脱帽蛋白结合使5′端帽子结构脱落, 然后被外切酶降解(Behm-Ansmant et al., 2006; Chen et al., 2009; Braun et al., 2011; Kuzuoğlu- Öeztüerk et al., 2012).miRNA还可以通过干扰翻译起始因子与5′端帽子结合、结合翻译抑制因子、促进核糖体与mRNA解离等方式抑制靶基因翻译(Nottrott et al., 2006; Mathonnet et al., 2007; Kiriakidou et al., 2007; Chendrimada et al., 2007; Fukaya et al., 2014).然而, 随着植物蛋白抗体研究的逐渐深入, 研究人员发现植物体内一些miRNA调控的靶基因蛋白表达水平下降, 但mRNA表达水平无明显变化(Auker-man and Sakai, 2003; Chen, 2004; Brodersen et al., 2008), 说明翻译抑制在植物miRNA中也存在.miR156/157可抑制靶基因SPL3翻译, 从而调控植物开花时间(Gandikota et al., 2007).研究人员通过放射性同位素标记, 发现拟南芥中miRNA存在抑制靶基因翻译现象且发生该过程的亚细胞位置在内质网(Li et al., 2013).植物AGO蛋白活性被抑制时miRNA无法剪切靶mRNA, 但对miRNA抑制靶基因翻译过程无显著影响(Iwakawa and Tomari, 2013), 说明植物miRNA介导靶mRNA剪切和抑制靶基因翻译的机制相对独立.但目前对miRNA选择作用模式的决定因素还不明确, 有待进一步研究. ... Nucleo-cytosolic shuttling of ARGONAUTE1 prompts a revised model of the plant microRNA pathway 1 2018 ... 植物miRNA在细胞核中合成(O'Brien et al., 2018; Bajczyk et al., 2023)(图1).高等植物中大多数miRNA基因代表独立的转录单元, 存在于基因间, 少数存在于蛋白质或非编码基因的内含子中(Millar and Waterhouse, 2005; Stepien et al., 2017; Pietrykowska et al., 2022).迄今为止, 已证实植物miRNA是由miRNA基因在RNA聚合酶II (RNA Pol-II)的作用下转录合成.在该酶的作用下, miRNA基因转录成具有5′端加帽和3′端多聚腺苷酸化的初级转录本(pri-miRNA), pri- miRNA折叠形成发夹状茎环结构(stem-loop).pri- miRNA被DCL1蛋白(DICER-LIKE1)或其它蛋白识别切割为前体miRNA (pre-miRNA), 并在双链RNA结合蛋白HYL1 (Hyponastic Leaves1)和锌指蛋白SE (Serrate)的作用下被进一步加工成miRNA/miRNA*复合体(Park et al., 2002; Lobbes et al., 2006; Dong et al., 2008; Bologna et al., 2009; Wang et al., 2019; Li and Yu, 2021; Xie et al., 2021).随后经甲基转移酶HEN1 (Hua Enhancer1)甲基化修饰3′端后与AGO蛋白结合, 去除miRNA*, 仅留下miRNA稳定在RNA介导的沉默复合物(RNA-induced silencing complex, RISC)中(Vaucheret et al., 2004; Yang et al., 2006; Baranauske et al., 2015; Tomassi et al., 2020; 武亮和戚益军, 2020).但miRNA从细胞核输出到细胞质中的过程仍不完全明确(Iki et al., 2010; Yu et al., 2017; Zhang et al., 2020a).miRNA与AGO蛋白结合后通过CRM1/EXPO1通路输出到细胞质中(Bologna et al., 2018), 然而也有部分研究发现miRNA存在以双链形式输出到细胞质中的可能性(Dalmadi et al., 2019).有研究表明, 与AGO蛋白结合输出到细胞质中的miRNA大多以转录后的方式形成, 而大部分以双链形式输出到细胞质中的miRNA以共转录的方式形成(Gonzalo et al., 2022). ... A loop-to-base processing mechanism underlies the biogenesis of plant microRNAs miR319 and miR159 1 2009 ... 植物miRNA在细胞核中合成(O'Brien et al., 2018; Bajczyk et al., 2023)(图1).高等植物中大多数miRNA基因代表独立的转录单元, 存在于基因间, 少数存在于蛋白质或非编码基因的内含子中(Millar and Waterhouse, 2005; Stepien et al., 2017; Pietrykowska et al., 2022).迄今为止, 已证实植物miRNA是由miRNA基因在RNA聚合酶II (RNA Pol-II)的作用下转录合成.在该酶的作用下, miRNA基因转录成具有5′端加帽和3′端多聚腺苷酸化的初级转录本(pri-miRNA), pri- miRNA折叠形成发夹状茎环结构(stem-loop).pri- miRNA被DCL1蛋白(DICER-LIKE1)或其它蛋白识别切割为前体miRNA (pre-miRNA), 并在双链RNA结合蛋白HYL1 (Hyponastic Leaves1)和锌指蛋白SE (Serrate)的作用下被进一步加工成miRNA/miRNA*复合体(Park et al., 2002; Lobbes et al., 2006; Dong et al., 2008; Bologna et al., 2009; Wang et al., 2019; Li and Yu, 2021; Xie et al., 2021).随后经甲基转移酶HEN1 (Hua Enhancer1)甲基化修饰3′端后与AGO蛋白结合, 去除miRNA*, 仅留下miRNA稳定在RNA介导的沉默复合物(RNA-induced silencing complex, RISC)中(Vaucheret et al., 2004; Yang et al., 2006; Baranauske et al., 2015; Tomassi et al., 2020; 武亮和戚益军, 2020).但miRNA从细胞核输出到细胞质中的过程仍不完全明确(Iki et al., 2010; Yu et al., 2017; Zhang et al., 2020a).miRNA与AGO蛋白结合后通过CRM1/EXPO1通路输出到细胞质中(Bologna et al., 2018), 然而也有部分研究发现miRNA存在以双链形式输出到细胞质中的可能性(Dalmadi et al., 2019).有研究表明, 与AGO蛋白结合输出到细胞质中的miRNA大多以转录后的方式形成, 而大部分以双链形式输出到细胞质中的miRNA以共转录的方式形成(Gonzalo et al., 2022). ... GW182 proteins directly recruit cytoplasmic deadenylase complexes to miRNA targets 1 2011 ... miRNA的另一种作用途径是抑制靶基因的翻译过程, 此途径在动物中较为常见, 在植物中的研究相对较少.动物miRNA仅能在种子序列区域(seed region)与靶基因3′端非编码区进行配对, 互补程度不高, 无法直接剪切mRNA, 因此抑制靶基因翻译是动物miRNA的常见作用模式(Huntzinger and Izaurralde, 2011).其作用机制为动物miRNA-RISC复合体除AGO蛋白外, 还含有GW182蛋白, 后者可以与翻译过程中众多因子相互作用.GW182蛋白可以与多腺苷酸结合蛋白相互作用, 使polyA在mRNA 3′端解离, 也可以与相关脱帽蛋白结合使5′端帽子结构脱落, 然后被外切酶降解(Behm-Ansmant et al., 2006; Chen et al., 2009; Braun et al., 2011; Kuzuoğlu- Öeztüerk et al., 2012).miRNA还可以通过干扰翻译起始因子与5′端帽子结合、结合翻译抑制因子、促进核糖体与mRNA解离等方式抑制靶基因翻译(Nottrott et al., 2006; Mathonnet et al., 2007; Kiriakidou et al., 2007; Chendrimada et al., 2007; Fukaya et al., 2014).然而, 随着植物蛋白抗体研究的逐渐深入, 研究人员发现植物体内一些miRNA调控的靶基因蛋白表达水平下降, 但mRNA表达水平无明显变化(Auker-man and Sakai, 2003; Chen, 2004; Brodersen et al., 2008), 说明翻译抑制在植物miRNA中也存在.miR156/157可抑制靶基因SPL3翻译, 从而调控植物开花时间(Gandikota et al., 2007).研究人员通过放射性同位素标记, 发现拟南芥中miRNA存在抑制靶基因翻译现象且发生该过程的亚细胞位置在内质网(Li et al., 2013).植物AGO蛋白活性被抑制时miRNA无法剪切靶mRNA, 但对miRNA抑制靶基因翻译过程无显著影响(Iwakawa and Tomari, 2013), 说明植物miRNA介导靶mRNA剪切和抑制靶基因翻译的机制相对独立.但目前对miRNA选择作用模式的决定因素还不明确, 有待进一步研究. ... Widespread translational inhibition by plant miRNAs and siRNAs 1 2008 ... miRNA的另一种作用途径是抑制靶基因的翻译过程, 此途径在动物中较为常见, 在植物中的研究相对较少.动物miRNA仅能在种子序列区域(seed region)与靶基因3′端非编码区进行配对, 互补程度不高, 无法直接剪切mRNA, 因此抑制靶基因翻译是动物miRNA的常见作用模式(Huntzinger and Izaurralde, 2011).其作用机制为动物miRNA-RISC复合体除AGO蛋白外, 还含有GW182蛋白, 后者可以与翻译过程中众多因子相互作用.GW182蛋白可以与多腺苷酸结合蛋白相互作用, 使polyA在mRNA 3′端解离, 也可以与相关脱帽蛋白结合使5′端帽子结构脱落, 然后被外切酶降解(Behm-Ansmant et al., 2006; Chen et al., 2009; Braun et al., 2011; Kuzuoğlu- Öeztüerk et al., 2012).miRNA还可以通过干扰翻译起始因子与5′端帽子结合、结合翻译抑制因子、促进核糖体与mRNA解离等方式抑制靶基因翻译(Nottrott et al., 2006; Mathonnet et al., 2007; Kiriakidou et al., 2007; Chendrimada et al., 2007; Fukaya et al., 2014).然而, 随着植物蛋白抗体研究的逐渐深入, 研究人员发现植物体内一些miRNA调控的靶基因蛋白表达水平下降, 但mRNA表达水平无明显变化(Auker-man and Sakai, 2003; Chen, 2004; Brodersen et al., 2008), 说明翻译抑制在植物miRNA中也存在.miR156/157可抑制靶基因SPL3翻译, 从而调控植物开花时间(Gandikota et al., 2007).研究人员通过放射性同位素标记, 发现拟南芥中miRNA存在抑制靶基因翻译现象且发生该过程的亚细胞位置在内质网(Li et al., 2013).植物AGO蛋白活性被抑制时miRNA无法剪切靶mRNA, 但对miRNA抑制靶基因翻译过程无显著影响(Iwakawa and Tomari, 2013), 说明植物miRNA介导靶mRNA剪切和抑制靶基因翻译的机制相对独立.但目前对miRNA选择作用模式的决定因素还不明确, 有待进一步研究. ... Genome-wide identification of GRF gene family and their contribution to abiotic stress response in pitaya (Hylocereus polyrhizus) 1 2022 ... 植物miR396-GRFs调控模块影响多个重要的生物学过程.双荧光素酶报告基因实验结果表明, ped-miR396d可以与PeGRF6同源物结合(Li et al., 2023a).构建miR396丰度降低和升高型拟南芥转化体, 发现miR396降低型植株鲜重和主根长等表型评估参数都得到增强, 而miR396升高型植株鲜重和主根长等均有所降低, 营养生长严重延迟, 施加盐胁迫处理后, 降低型拟南芥光合色素含量相比野生型和miR396升高型植株更高, 对盐胁迫耐受性更强(Pegler et al., 2021a).缺氮条件下, 水稻miR396敲除型植株地上部生物量相较野生型增加25%, 氮转运和代谢相关基因表达上调, miR396的敲除导致植株氮利用率提高, 与其靶基因OsGRF4过表达植株表型一致, 说明miR396调节植物氮同化及氮代谢(Zhang et al., 2020b), 低磷条件下也有相似的实验结果(Pegler et al., 2021b).此外, 众多研究表明, miR396在火龙果(Hylocereus polyrhizus)、菊花(Chrysanthemum indicum)、凤丹(Paeonia ostii)、番茄、苜蓿、花生(Arachis hypogaea)和豇豆(Vigna unguiculata)等多个物种中均能通过促进水分及营养元素吸收、增加光合色素含量和增加籽粒产量和质量等途径增强植物对非生物胁迫的耐受性(Pegler et al., 2020; Fracasso et al., 2021; Liu et al., 2021; Cai et al., 2022; Guo et al., 2022a; Zhang et al., 2022; Hasan et al., 2023; Ding et al., 2024). ... GmTIR1/GmAFB3-based auxin perception regulated by miR393 modulates soybean nodulation 1 2017 ... miR393靶向F-box生长素受体TIR1和生长素信号转导蛋白AFBs基因, 通过释放生长素反应因子(ARF)和介导Aux/IAA阻遏蛋白水解调节生长素信号转导以响应非生物胁迫(Chen et al., 2011; Wojcik and Gaj, 2016; Cai et al., 2017).冷胁迫处理后植物冷响应基因表达增强, 过表达miR393植株相关基因表达量增幅更高, 对低温耐受性更强, 外源生长素处理负调控上述冷响应基因, 说明生长素信号通路参与植物耐寒性, 这表明miR393抑制的生长素信号转导能增强植物的抗寒性(Liu et al., 2017).对铝胁迫下的大麦施加外源生长素可增强Al3+诱导的根系抑制作用, 且Al3+诱导根尖生长素应答基因表达, miR393的过表达可减弱这种抑制作用并下调相关反应基因的表达, 表明miR393通过改变大麦生长素信号输出调控根系对Al的敏感性(Bai et al., 2017).研究发现, miR393调节生长素反应因子进而调控气孔发育相关基因的表达, 过表达植株表现出叶片保卫细胞长度变短, 气孔密度增加, 植株对干旱胁迫更敏感, 而敲除系植株则呈现相反的表型(Yuan et al., 2019b).在匍匐翦股颖和大豆(Glycine max)等植物中也获得了相似的结果(Zhao et al., 2019; Xing et al., 2023).也有研究表明, miR393通过盐胁迫诱导的应激激素脱落酸与生长素之间的拮抗作用参与植物对盐胁迫的响应(Denver and Ullah, 2019). ... Small RNA and degradome deep sequencing reveals drought- and tissue-specific micrornas and their important roles in drought-sensitive and drought-tolerant tomato genotypes 2 2016 ... 随着微阵列和高通量深度测序的快速发展, 研究人员在植物响应非生物胁迫方面鉴定出大量miRNA.对不同植物进行干旱、温度和盐等胁迫处理后鉴定响应相关胁迫的miRNA (表1).研究发现, 对不同品种番茄(Solanum lycopersicum)幼苗进行干旱胁迫处理后, 分别以根系和地上部为材料建立小RNA文库进行深度测序, 鉴定出来自578个家族的699个miRNAs, 其中干旱耐受型品种在根系中差异表达的miRNA为101个, 在地上部中差异表达的miRNA为58个, 干旱敏感型品种在根系和地上部中差异表达的miRNA分别为46和61个(Candar-Cakir et al., 2016).拟南芥幼苗在干旱、高温和盐胁迫处理下相较对照组差异表达的miRNA分别为123、121和118个, 包含特异性响应干旱胁迫miRNA 33个、特异性响应盐胁迫miRNA 25个、特异性响应高温胁迫miRNA 24个以及在三者中共同响应的miRNA 52个(Pegler et al., 2019).这表明非生物胁迫下植物miRNA的鉴定结果受物种、品种、胁迫处理方法、处理时间及组织特异性等因素影响. ...
本文的其它图/表
|

 首页
首页