
|
|
||
|
高温胁迫影响玉米生长发育的分子机制研究进展
植物学报
2024, 59 (6):
963-977.
DOI: 10.11983/CBB24049
植物在其生命周期中面临多种非生物胁迫, 如高温、干旱和盐碱, 这些胁迫对植物的生长发育产生多种影响。全球变暖加剧了高温胁迫对玉米(Zea mays)等作物的影响, 可能导致其生长受阻和生殖能力下降。玉米作为重要的农作物, 其产量和品质受高温胁迫影响严重。植物通过复杂的分子机制响应高温, 涉及多个信号转导途径和基因表达调控。利用遗传学、基因组学、多组学分析以及高通量表型分析等前沿技术, 深入挖掘和解析玉米基因组中非生物胁迫耐受性(包括热胁迫)的相关基因和位点至关重要。这些研究不仅有助于深入理解玉米耐受胁迫的生物学机制, 而且为加快玉米新品种培育提供了关键的分子标记和候选基因资源。 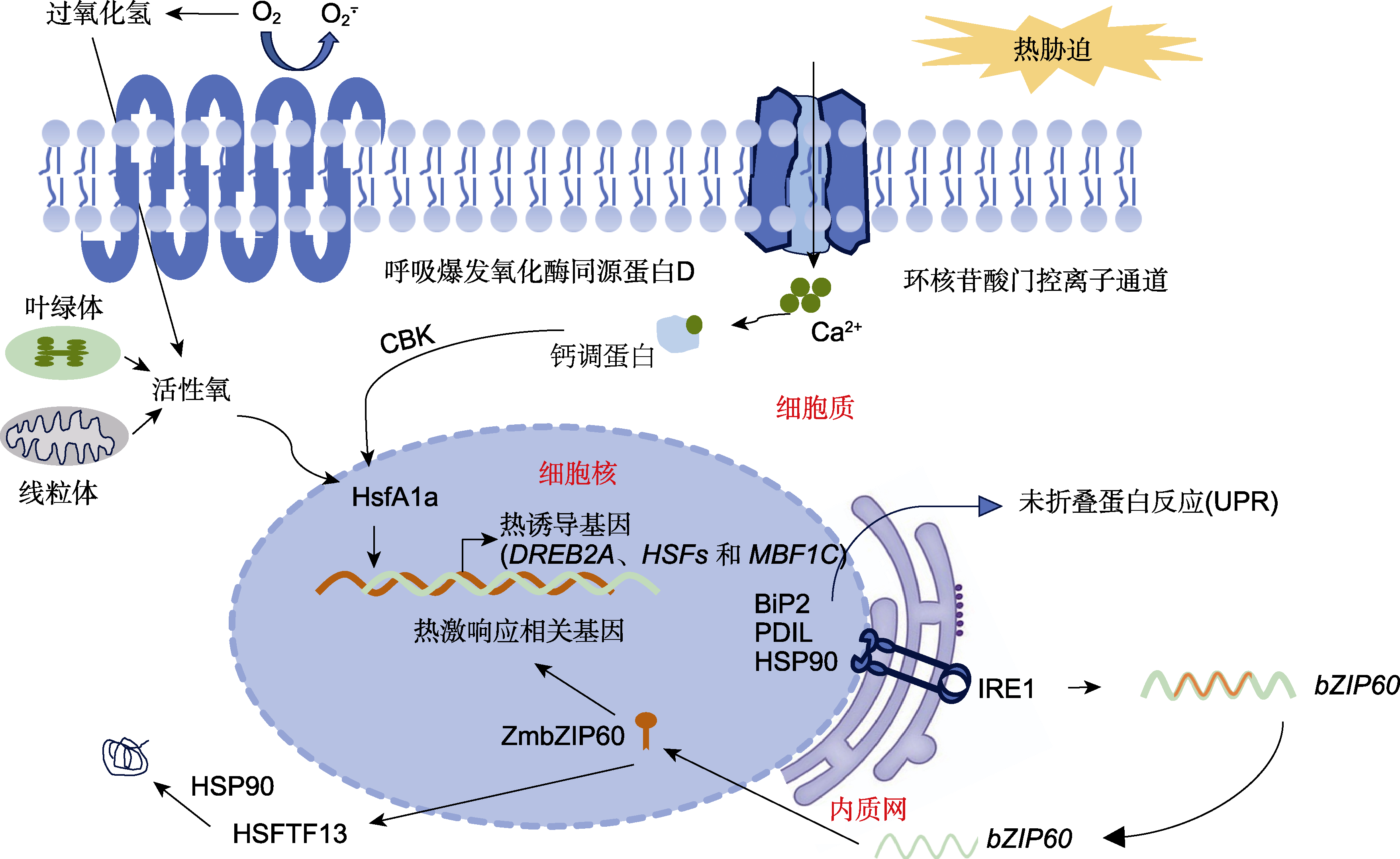 View image in article
图1
植物响应高温胁迫的调控网络
当植物暴露于高温胁迫下, 其细胞膜受损伤, 质外体Ca2+释放, 导致叶绿体和线粒体功能丧失, 细胞质内Ca2+和活性氧(ROS)水平升高, 以及错误折叠或未折叠蛋白质过度积累。细胞质内Ca2+和ROS增加并作为第二信使刺激下游调控网络。高温胁迫破坏蛋白质稳态, 诱导未折叠蛋白反应(UPR)以及由IRE1-ZmZIP60介导的信号通路。ZmZIP60激活A型热激转录因子(HSFs)和HSFTF13的表达, 后者上调热激蛋白(HSP)基因(如HSP90)的表达。
正文中引用本图/表的段落
钙离子依赖的蛋白激酶(calcium-dependent protein kinases, CDPKs)能够直接结合钙离子并磷酸化下游底物蛋白, 进而激活下游信号转导。在玉米中已鉴定出40个CDPKs。高温胁迫下, 活化的CDPKs磷酸化热激转录因子(HSFs), 后者进入细胞核, 并启动HSPs (如HSP40、HSP70、HSP90和HSP100)的表达(Qian et al., 2019)。其中ZmCDPK7位于细胞质膜上, 受高温正调控。在高温胁迫下, ZmCDPK7从细胞质膜转移到细胞质, 并磷酸化下游底物蛋白, 从而提高玉米的耐热性(Zhao et al., 2021)。钙调蛋白结合转录激活因子(calmodulin-binding transcription activator, CAMTA)是存在于真核生物中结构保守的重要钙调素结合蛋白, 广泛参与植物的胁迫应答(图1)。研究发现, 玉米ZmCAMTA1、ZmCAMTA2和ZmCAMTA3在高温胁迫下显著上调表达(Yue et al., 2015; El-Sappah et al., 2022)。
高温胁迫导致植物体内活性氧类物质积累, 包括过氧化氢(hydrogen peroxide, H2O2)、羟基自由基(·OH)和超氧自由基(O2-. )等(图1)。ROS作为关键的次级信号分子, 在植物响应高温胁迫过程中扮演着重要角色(Sewelam et al., 2014)。正常情况下, 细胞通过自身的抗氧化系统确保ROS的产生和清除处于稳定的平衡状态, 从而维持细胞内ROS水平在较低范围内。然而, 当植物受到高温胁迫时, ROS的产生显著增加, 使原本平衡的ROS产生与清除机制被打破。因此, 植物细胞内ROS逐渐积累, 导致氧化损伤。如果ROS含量过高, 将对细胞造成严重损伤, 甚至导致细胞死亡。一旦遭遇高温胁迫, ROS分子便迅速生成, 并触发植物的高温胁迫响应机制(Volkov et al., 2006; Mittler et al., 2012)。值得注意的是, 当用H2O2处理植物时, 其耐热性显著提升。然而, 如果编码生成ROS的RBOHs基因(如AtRBOHB和AtRBOHD)发生突变, 那么植物在应对高温胁迫时其耐热性就会受到抑制(Larkindale and Huang, 2004; Larkindale et al., 2005)。为应对高温带来的影响, 玉米通过激活MYB、AP2/EREBP、NAC、BRs、HSPs、Rubisco、抗氧化剂(如抗坏血酸过氧化物酶和谷胱甘肽S-转移酶)以及激酶等基因清除过多的活性氧(ul Haq et al., 2019; Jagtap et al., 2020)。一方面, H2O2直接促进HSFA1a、HSFA4a和HSFA8转录因子的激活; 另一方面, H2O2与一氧化氮(NO)信号共同作用, 直接激活HSFA1α、HSFA4α和HSFA8, 从而诱导玉米热响应基因的表达。这些反应诱导热激响应元件(heat shock response element, HSE)与热休克蛋白(heat shock proteins, HSPs)启动子结合(Miller and Mittler, 2006; Li et al., 2018; El-Sappah et al., 2022)。
在高温胁迫下植物会产生热激反应(heat shock response, HSR), 热激蛋白在此过程中发挥重要作用。在应对高温胁迫的过程中, 热激蛋白通过与蛋白质疏水表面结合, 在维持蛋白质功能构象、防止非天然蛋白聚集、促进蛋白重新折叠并组装以及降解蛋白质等过程中发挥作用。根据分子量的大小, HSPs可以分为6类, 即HSP100、HSP90、HSP70、HSP60、HSP40和小分子热激蛋白(sHSP)。热激蛋白具有分子伴侣的功能, 分子伴侣是指参与新生肽链的折叠、功能性蛋白质或蛋白质复合体组装及转运的一类蛋白质, 该类蛋白质不参与蛋白质的合成过程, 且蛋白质组装完成后便解离出来。在常温下, HSP70/HSP90等结合HSFA1s, 阻止其与热激响应元件(HSE)结合, 使其无法诱导热响应基因包括HSPs的表达(图1)。高温胁迫下, 热激反应途径被激活, 诱导HSP70/HSP90与热激转录因子解离, 热激转录因子活化形成三聚体, 与热激蛋白上游启动子中的HSE结合, 迅速诱导热激蛋白的表达(Ohama et al., 2017)。热激蛋白可阻止细胞内蛋白质发生不可逆变性, 有助于细胞增强对高温的耐受性。部分热激蛋白在响应胁迫时具有组织特异性。玉米幼苗暴露于高温后, ZmHSPs的表达量增高数百倍甚至数千倍。其中, ZmHSP100s、ZmHSP90s、ZmHSP70s、ZmCPN60s、ZmHSP40s以及ZmsHSPs均表现出强烈的表达升高, 增幅最高可达3 000倍(Diogo et al., 2023)。与营养部位(如叶和根)相比, HSP101在玉米生殖部位(如雄穗、穗和胚乳)的表达增高更显著。这表明HSP101在玉米的生殖阶段可能发挥更为关键的作用(Young et al., 2001)。
热激转录因子处于HSR信号通路的关键位置, 在植物抵抗高温胁迫过程中发挥至关重要的作用(Timperio et al., 2008)。高温胁迫能够迅速诱导多种热休克响应转录因子的表达, 包括脱水响应元件结合蛋白2A (DREB2A)、HSFs以及多蛋白结合因子(MBF1C) (图1) (Zhao et al., 2021)。热激转录因子是植物响应高温胁迫的重要调控因子。HSFs分为A、B和C三类。在玉米中, 已报道了31个HSFs, 其中一些属于A类的热激转录因子被证实与高温耐受性有关(Lin et al., 2011; Zhang et al., 2020)。HSFA1s在高温胁迫响应中发挥关键作用, 对转录网络的激活起主要调节作用(Liu et al., 2011; Andrási et al., 2021)。HSFA1直接参与调控热激反应重要基因(HSFA7s、HSFA2、HSFBs、MBF1c和DREB2A)的表达, 进而调节伴侣蛋白和酶的合成, 促进未折叠蛋白的折叠, 降解错误折叠蛋白。同时, 高温胁迫还促进伴侣蛋白HSP70与HSP90之间的相互作用, 进一步激活HSFA1 (Ohama et al., 2017)。ZmHsf-6属于A1类, ZmHsf-1、ZmHsf-4、ZmHsf-5和ZmHsf-17属于A2类, 而ZmHsf-3、ZmHsf-11和ZmHsf-25属于B类, 这些热休克因子在玉米耐热性中均扮演关键角色(Lin et al., 2011; Zhang et al., 2020; Jiang et al., 2021)。ZmHsf-6受多种逆境胁迫, 尤其是高温胁迫诱导表达显著上调, 过表达ZmHsf-6拟南芥植株的耐热性明显提高(Li et al., 2015a)。同时, 检测发现ZmHsf-6在花粉中表达, 且在高温胁迫下其表达水平上调(Jiang et al., 2021)。而在高温胁迫下的玉米植株中, ZmHsf-1、ZmHsf-3和ZmHsf-23的表达量最高, 这表明它们在增强玉米耐热性方面具有重要作用(Lin et al., 2011)。最新研究发现, 通过热胁迫转录组图谱和共表达网络分析, 发现核心热激转录因子ZmHSF20在玉米响应热胁迫过程中发挥重要作用。ZmHSF20对纤维素合成酶基因ZmCesA2和热激转录因子ZmHsf4的表达均具有抑制作用, 而ZmHSF4与ZmCesA2具有转录激活表达的调控关系。高温胁迫下, ZmHsf20过表达株系对高温更敏感, 而Zmhsf20突变体耐热性增强, 这表明ZmHSF20负调控玉米的耐热性(Li et al., 2024)。
高温胁迫下, 错误折叠蛋白促进热激蛋白HSP70家族成员BIP的富集, BIP随后从内质网膜结合蛋白IRE1上解离下来。BIP在错误折叠蛋白与IRE1之间起桥梁作用, 促使其形成激活型二聚体。激活的IRE1能够剪接基本亮氨酸拉链结构域(ZmbZIP60)的mRNA。经过剪接的ZmbZIP60 mRNA被导入细胞核, 进而编码专门响应高温胁迫的ZmbZIP60转录因子。在玉米中, 当IRE1被激活时, 其能够将ZmbZIP60转录本剪接为特定的mRNA形式, bZIP60s能够激活A型热休克因子Hsftf13, 后者进一步激活一系列热休克蛋白(HSP)基因的表达(图1) (Korner et al., 2015; Li et al., 2020b)。
本文的其它图/表
|
 首页
首页{kind=link}