
|
|
||
|
lncRNA调控玉米生长发育和非生物胁迫研究进展
植物学报
2024, 59 (6):
950-962.
DOI: 10.11983/CBB24075
长链非编码RNA (long non-coding RNA, lncRNA)广泛存在于真核生物基因组中, 在维持生物体正常生命活动中发挥重要作用。近年来, 通过高通量测序和生物信息学分析在植物中发掘到大量的lncRNA。已有研究证实lncRNA在调控植物生长发育和逆境响应中发挥重要作用。由于基因组复杂且遗传操作过程繁琐, lncRNA在玉米(Zea mays)中的研究远落后于拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)。玉米作为我国主要粮食作物, 对于保障国家粮食安全至关重要。玉米还是遗传学与基因组学领域重要的模式植物。了解lncRNA在玉米中的研究进展有助于理解lncRNA的生物学功能。挖掘并解析lncRNA参与玉米生长发育和逆境响应的分子调控网络, 可为玉米遗传改良提供新的分子靶点。该文总结了lncRNA的来源、分类和作用机制, 并讨论了玉米中lncRNA的发掘及其在调控生长发育和逆境响应中的生物学功能, 最后展望了lncRNA在玉米中的研究方向。  View image in article
图1
长链非编码RNA (lncRNA)的分类
蓝色框代表mRNA的外显子, 红色线表示lncRNA转录本, 黑色箭头表示转录方向。
正文中引用本图/表的段落
为方便学术成果分享, 对lncRNA的分类和功能注释至关重要。根据lncRNA在基因组中与邻近编码蛋白基因的位置关系, 可将lncRNA分为基因间lncRNA (intergenic lncRNA, lincRNA)、正义链lncRNA (sense lncRNA)、天然反义链lncRNA (natural antisense lncRNA, NATs)、内含子lncRNA (intronic lncRNA)和双向lncRNA (bidirectional lncRNA) (图1) (Hermans-Beijnsberger et al., 2018)。目前已知的植物功能性lncRNA类型主要为lincRNA (如PROMPTs、HID1和MISSEN)、NAT (如MAS、COOLAIR、cis- NATPHO1;2及LAIR)和内含子lncRNA (如COLDAIR) (Heo and Sung, 2011; Jabnoune et al., 2013; Csorba et al., 2014; Wang et al., 2014, 2018; Kim and Sung, 2017; Zhao et al., 2018; Song et al., 2019; Zhou et al., 2021)。正义链lncRNA与双向lncRNA是否参与植物生长发育和逆境胁迫响应还有待进一步证实。
在动物中lncRNA作为信号分子、诱饵分子、引导分子和支架分子等, 通过顺式(cis)和反式(trans)作用调控靶标基因的表达(Wang and Chang, Origins and evolving functionalities of tRNA-derived small RNAs 1 2021 ... 真核生物基因组中高达90%的区域可被转录, 这些转录产物大部分为非编码RNA (non-coding RNA, ncRNA).人类基因组中仅有不到2%的转录产物具有蛋白编码功能(Djebali et al., Plant and animal small RNA communications between cells and organisms 1 2022 ... 真核生物基因组中高达90%的区域可被转录, 这些转录产物大部分为非编码RNA (non-coding RNA, ncRNA).人类基因组中仅有不到2%的转录产物具有蛋白编码功能(Djebali et al., Regulation of phosphate homeostasis by microRNA in Arabidopsis 1 2006 ... 植物缺磷是全球农业生产中普遍面临的问题.深入理解植物适应磷胁迫的生理及分子机制, 充分挖掘作物自身吸收和利用磷素的生物学潜力, 有助于实现磷肥的高效利用.miRNA399是第1个被发现响应低磷胁迫的植物miRNA, 其特异性受低磷胁迫诱导表达(Fujii et al., Antisense COOLAIR mediates the coordinated switching of chromatin states at FLC during vernalization 2 2014 ... 为方便学术成果分享, 对lncRNA的分类和功能注释至关重要.根据lncRNA在基因组中与邻近编码蛋白基因的位置关系, 可将lncRNA分为基因间lncRNA (intergenic lncRNA, lincRNA)、正义链lncRNA (sense lncRNA)、天然反义链lncRNA (natural antisense lncRNA, NATs)、内含子lncRNA (intronic lncRNA)和双向lncRNA (bidirectional lncRNA) (
在动物中lncRNA作为信号分子、诱饵分子、引导分子和支架分子等, 通过顺式(cis)和反式(trans)作用调控靶标基因的表达(Wang and Chang, Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA 2 2011 ... 为方便学术成果分享, 对lncRNA的分类和功能注释至关重要.根据lncRNA在基因组中与邻近编码蛋白基因的位置关系, 可将lncRNA分为基因间lncRNA (intergenic lncRNA, lincRNA)、正义链lncRNA (sense lncRNA)、天然反义链lncRNA (natural antisense lncRNA, NATs)、内含子lncRNA (intronic lncRNA)和双向lncRNA (bidirectional lncRNA) (
在动物中lncRNA作为信号分子、诱饵分子、引导分子和支架分子等, 通过顺式(cis)和反式(trans)作用调控靶标基因的表达(Wang and Chang, Comparison of stranded and non-stranded RNA-seq transcriptome profiling and investigation of gene overlap 1 2015 ... 在构建链特异性转录组测序(strand-specific RNA sequencing, ssRNA-seq)文库时, 将转录本的方向信息保存到测序文库中, 能够确定转录本来源于基因组的正链或负链.因此, ssRNA-seq可用于几乎所有类型的lncRNA挖掘, 并且对转录本的表达定量更加准确(Zhao et al., Global identification of Arabidopsis lncRNAs reveals the regulation of MAF4 by a natural antisense RNA 2 2018 ... 为方便学术成果分享, 对lncRNA的分类和功能注释至关重要.根据lncRNA在基因组中与邻近编码蛋白基因的位置关系, 可将lncRNA分为基因间lncRNA (intergenic lncRNA, lincRNA)、正义链lncRNA (sense lncRNA)、天然反义链lncRNA (natural antisense lncRNA, NATs)、内含子lncRNA (intronic lncRNA)和双向lncRNA (bidirectional lncRNA) (
本文的其它图/表
|
 首页
首页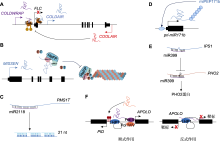
{kind=link}