|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
广义蕨类植物基因功能研究概述
植物学报
2024, 59 (3):
495-514.
DOI: 10.11983/CBB23090
广义蕨类植物(石松类和蕨类)是陆生维管植物第二大类群, 兼具孢子植物和维管植物的结构和生理特点, 衔接陆生植物从简单到复杂的演化过程。广义蕨类植物表现出许多不同于其它植物类群的独特生物学过程, 在研究植物基因组演化、器官发育、繁殖方式及环境适应等方面是一类关键的植物类群。现代测序技术的快速发展和应用推动了广义蕨类植物全基因组测序和组装, 为其基因功能研究提供了巨大便利。为充分认识广义蕨类植物基因功能研究的热点和难点, 该文从多角度全面总结了广义蕨类植物基因功能研究, 重点论述广义蕨类植物在器官发育、生殖过程、环境适应性和次生代谢物合成等方面的研究现状, 系统阐述其在揭示广义蕨类植物特殊生物学过程中的应用及各类基因的功能。最后, 对如何加快广义蕨类植物基因功能研究, 并利用相关研究揭示其生物学特性和开发更广泛的应用场景提出了建议。
表1
已发表全基因组序列的广义蕨类植物
正文中引用本图/表的段落
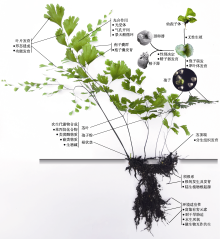
解析植物全基因组序列是从遗传的全局角度理解基因的功能及数目演化、新基因的起源、通路相关基因的出现或丢失等关键生物演化事件, 而蕨类植物是研究基因演化过程的重要节点类群。与被子植物相比, 蕨类植物的基因组普遍较大(Wang et al., 2022), 且染色体数目多, 导致该类群物种染色体级别的全基因组测序和组装难度较高(图2)。截至2023年10月, 共有8种石松类和6种蕨类物种完成全基因组测序和组装(Marks et al., 2021; Sun et al., 2022) (
目前, 有关陆生维管植物叶起源的解释有2种假说: 突起学说和顶枝学说(Wilson, 1953)。突起学说认为石松类植物的小型叶起源于茎轴表面的突起体, 而顶枝学说认为大型叶和小型叶都是由原始复合顶枝的次要二叉分枝经耸出、联合及扁化发展而来。蕨类植物的器官结构简单, 但叶片形态丰富, 其复杂程度不低于被子植物, 是研究植物叶片起源和形态建成的理想材料。新的分子证据表明, 植物小型叶和大型叶的形成可能由相同的发育机制调控。通过研究KNOX (KNOTTED1-LIKE HOMEOBOX)和ARP (ASYMMETRIC LEAVES1, ROUGH SHEATH2 and PHANTASTICA)在翠云草(Selaginalla uncinata)和欧紫萁(Osmunda regalis)分生组织中的表达, 发现KNOX和ARP在茎顶端分生组织共表达促进分枝产生, 在叶原基共表达导致出现复杂叶(Harrison et al., 2005)。在拟南芥中分别过表达美洲水蕨CrKNOX1和CrKNOX2, 所得转基因拟南芥的叶片形态均发生显著改变, 叶片边缘出现类似美洲水蕨叶片的深度裂刻(Sano et al., 2005)。野胡萝卜(Daucus carota)、白粉藤(Cissus congestusm)、佛罗里达泽米(Zamia floridans)以及翠蕨(Anogramma chaeophylla)的叶片均具有复杂结构, 通过比较植物KNOXI (I类)在叶原基的表达模式, 研究者认为蕨类植物和被子植物复杂叶片经历相对独立的演化历程(Bharathan et al., 2002)。同样地, 通过比较舌蕨属(Elaphoglossum spp.) 3个不同种E. lloense (简单叶)、E. peltatum f. standleyi (简单叶)和E. peltatum f. peltatum (复杂叶) KNOXI在叶原基的表达模式, 发现叶原基裂刻中KNOXI的持续表达与叶片复杂结构的形成有关, 但KNOXI不是决定叶片复杂结构形成的唯一遗传因素(Vasco and Ambrose, 2020)。Cruz等(2020)在Mickelia scandens中鉴定到3个KNOX基因, 其中MsC1KNOX1和MsC1KNOX2在茎顶端分生组织、原形成层以及叶顶端分生组织和间叶分生组织中表达, 参与叶片发育。综上所述, KNOX在调节蕨类植物叶片形态发育方面具有保守性, 与其它遗传因子共同调节叶片形态建成。
脱落酸(ABA)参与植物的配子体发育。SnRK2 (SNF1-related kinase2)是ABA信号通路的核心调节因子。为探明ABA-SnRK2信号通路在蕨类植物中的起源与演化过程, McAdam等(2016)通过筛选对ABA不敏感的美洲水蕨突变体, 鉴定到1个SnRK2的同源基因CrGAIA1; 功能验证表明ABA-CrGAIA1信号通路参与美洲水蕨配子体性别分化和孢子萌发, 这与ABA-SnRK2信号通路在种子植物中的功能类似, 不同的是ABA-CrGAIA1不能调节美洲水蕨叶片气孔运动。铁线蕨AtLEC1及相关基因AdcFUS3和AdcABI3在同型孢子囊中共表达, 该分子模块可能通过同化物积累参与同型孢子囊的发育过程(Fang et al., 2022)。GA与ABA在调节种子植物一些生物学过程时存在功能拮抗现象, 在蕨类植物中二者如何调节配子体性别分化有待深入研究。
种子植物尤其是高大乔木大量出现后迅速占领地球陆地部分的上层生存空间, 矮小的蕨类植物只能与其它灌木一起竞争林下荫蔽生存空间。为适应少光黯淡的生境, 部分蕨类植物演化出特有的适应机制。Kawai等(2003)发现铁线蕨光受体突变体rap (phy3)的叶绿体对蓝光和暗白光敏感性降低, 失去原有的向光性。研究表明, 铁线蕨具有被子植物常见的光受体PHOT1和PHOT2, 但phy3结构异于上述光受体, 且普遍存在于水龙骨目蕨类植物Dryopteris filix-mas、Onoclea sensibilis和Hypolepis punctata中, 在简单蕨类(欧紫萁和海金沙)和种子植物中未发现phy3的同源基因。研究者认为phy3的起源是真叶蕨类在以种子植物为主的荫蔽森林环境下逆势种群激增的关键遗传条件(Kawai et al., 2003)。
Yan等(2019)总结有6类蛋白参与蜈蚣草中砷化合物的转运, 包括ACR3 (arsenical compound resistance 3)、ABC亚家族、P-type ATPase、MFS亚家族(major facilitator superfamily)、MIP (major intrinsic protein)和NRT3亚家族(nitrate transporter 3)。PvACR3是定位在液泡膜上的砷酸盐转运蛋白, 受砷胁迫诱导表达; 它将砷酸盐转运至液泡内并区隔化, 保护细胞免受砷毒害(Indriolo et al., 2010)。在拟南芥中异源过表达PvACR3能显著提高转基因拟南芥的砷耐受性和砷富集(Chen et al., 2013)。但该基因在被子植物中丢失, 这可能是被子植物普遍不耐砷胁迫的原因之一。PvACR3的另外2个同源基因PvACR3;2和PvACR3; 3分别定位于质膜和液泡膜, 负责亚砷酸盐的长距离运输和区隔化(Chen et al., 2021)。与蜈蚣草近缘的凤尾科(Pteridaceae)植物凤尾蕨(P. cretica) PcACR3基因与PvACR3功能类似(Popov et al., 2021)。水通道蛋白PvTIP4;1定位于质膜, 在蜈蚣草根部特异性表达, 异源表达后促进拟南芥根系对亚砷酸盐的吸收(He et al., 2016)。Pht1 (phosphate transporter 1)是一类磷酸盐转运蛋白。DiTusa等(2016)在蜈蚣草中鉴定到3个PvPht1基因, 分别是PvPht1;1、PvPht1;2和PvPht1;3。其中PvPht1;3基因表达响应磷亏缺和砷酸盐胁迫, 有高砷酸盐亲和性, 能使酵母积累更多的砷酸盐。PvPht1;3可能是蜈蚣草具备超砷富集能力的关键因子。Feng等(2021)在蜈蚣草中鉴定到磷酸盐转运蛋白PvPht2;1, PvPht2;1基因定位于叶绿体被膜, 在茎叶中高表达, 但不响应磷亏缺和砷胁迫; 砷胁迫条件下PvPht2;1过表达拟南芥未富集砷化物, 但地上部茎叶和叶绿体中的磷元素含量升高; 研究发现PvPht2;1促进磷元素进入叶绿体保护植物的光合系统, 以减轻砷毒害。
次生代谢物是维管植物具备复杂结构和环境适应性的物质基础。相较于最古老的维管植物, 江南卷柏基因组中参与次生代谢物合成的基因数量显著增多, 如细胞色素P450单加氧酶(cytochrome P450, CYP450)、酰基转移酶和萜类合酶(Banks et al., 2011)。这些酶参与苯丙素类、萜类、生物碱、甾体及鞣质等次生代谢物的合成。比较代谢组学研究表明, 美洲水蕨中有许多被子植物中无或作用未知的特异性次生代谢物(Marchant et al., 2022), 这些物质的生物活性和代谢途径有待深入研究。目前, 蕨类植物次生代谢途径相关基因的研究多限于基因鉴定、克隆和序列分析等分子研究的初级阶段, 仅有少数基因通过异源遗传转化技术进行功能验证, 几乎未见同源功能验证的案例(表1">附
体外酶活实验表明, 种子植物CYP98 (cytochrome P450 monooxygenase 98)的底物种类丰富, 4-香豆酰莽草酸酯是裸子植物火炬松(Pinus taeda) PtCYP98的最优催化底物; 但是江南卷柏和蜈蚣草CYP98s对底物4-香豆酰莽草酸酯催化效率低, 更易催化4-香豆酰邻氨基苯甲酸酯; 被子植物CYP98催化底物丰富多样, 为其适应复杂多变的生存环境提供了更多保护性物质(Alber et al., 2019)。咖啡酰基辅酶A-3-O-甲基转移酶(CCoAOMTs)是木质素合成的关键酶, 友水龙骨(Goniophlebium amoenum)含有2个CCoAOMTs, 原核表达的GaCCoAOMT1和GaCCoAOMT2蛋白能甲基化底物咖啡酰基辅酶A, 催化效率与紫花苜蓿(Medicago sativa) MsCCoAOMT相近(Zhang et al., 2019)。
蕨类植物是地球上较早出现的陆生维管植物, 是研究多成员家族基因起源与演化的关键植物类群; 绿色植物由简单到复杂的演化过程中, 不同物种基因组中家族基因的种类、数目和功能会发生改变。基因组测序显示美洲水蕨MADS-box基因家族有35个成员, 但蕨类植物演化至美洲水蕨这一分支出现时一些特异的MADS- box转录因子丢失(Marchant?? et al., 2022)。LFY (LEAFY)转录因子基因是调节植物花器官起始的关键基因(Siriwardana and Lamb, 2012)。Rodríguez-Pelayo等(2022)构建了蕨类植物LFY基因的系统进化树, 发现一些特异的基因重复事件, 为研究LFY基因的分化提供了新思路。LFY基因在楔叶铁线蕨(A. raddianum)、巨木贼(Euisetum giganteum)和江南卷柏的营养叶和生殖叶各发育时期表达, 尤其在早期生殖发育和孢子囊分化时期表达量很高, 说明LFY基因已参与蕨类植物的生殖结构发育(Rodríguez- Pelayo et al., 2022)。KNOX转录因子调节植物的形态发育, 通过分析早期非维管植物类群轮藻(charophytes)、苔类(liverworts)、藓类(mosses)、角苔(hornworts)与维管植物KNOX基因的系统演化关系, 推测KNOX基因的分化(I类和II类)发生在植物登陆过程中。突变体基因功能回补实验证实, 藓类MKN2蛋白比维管植物KNOX蛋白能在更广阔的背景下行使功能(Frangedakis et al., 2017)。KANADI是GARP家族转录因子基因, 其DNA结合域序列在蕨类植物和种子植物中较为保守。原位杂交实验表明, 木贼(E. hyemale) EqhKAN1/2/3基因在远轴端分生组织中表达, 与种子植物类似; 但卷柏叶中SmKAN1/2/3基因无极性表达, 与其它蕨类植物和种子植物不同(Zumajo- Cardona et al., 2019)。
附表1 广义蕨类植物基因功能研究概况
- 未知 ... The telome theory 1 1953 ... 目前, 有关陆生维管植物叶起源的解释有2种假说: 突起学说和顶枝学说(Wilson, Differential gene expression profiling of one- and two-dimensional apogamous gametophytes of the fern Dryopteris affinis ssp 1 2020 ... 蕨类植物孢子萌发后由单个细胞发育成单层多细胞的原叶体, 此发育过程为研究植物细胞极性生长和器官发育提供了天然材料.通过比较Dryopteris affinis ssp. affinis一维(丝状体)到二维(原叶体)的转录组时空变化, 研究人员鉴定到一些差异表达基因, 参与分生组织发育、生殖过程、生长素信号转导和糖代谢等生物学过程(Wyder et al., Identification and characterization of L-lysine decarboxylase from Huperzia serrata and its role in the metabolic pathway of lycopodium alkaloid 1 2017 ... 生物碱是植物体内一类含氮杂环有机化合物.石松碱(即石松类植物生物碱的统称) (Siengalewicz et al., Rapid reduction of arsenate in the medium mediated by plant roots 1 2007 ... 蜈蚣草根系从根际环境吸收2种砷化物: 砷酸盐和亚砷酸盐(Ma et al., Genome analysis of the ancient tracheophyte Selaginella tamariscina reveals evolutionary features relevant to the acquisition of desiccation tolerance 1 2018 ... Pteridophytes with whole-genome sequence published
- 未知 ... Insecticidal fern protein Tma12 is possibly a lytic polysaccharide monooxygenase 1 2019 ... 在应用基础研究领域, 蕨类植物是中药活性成分开发(马尾杉属、金毛狗(Cibotium barometz)、槲蕨属(Drynaria))、环境污染治理(蜈蚣草)、城市景观绿化(巢蕨属(Asplenium)、桫椤科(Cyatheaceae)、鹿角蕨属(Platycerium))、生态修复(卤蕨属(Acrostichum))和发展林下经济(药用、食用和观赏蕨类)的重要植物资源.蕨类植物历经3亿年的演化, 我们对其在环境适应方面的分子机制知之甚少, 蕨类植物合成的具有生物活性的次生代谢物不胜枚举, 但人类对蕨类植物的认识和利用十分有限, 揭示其生物学特性的分子机制将为开发蕨类资源应用场景提供理论支撑.宽叶铁线蕨(A. latifolium)中的萜类化合物(22-hydroxyhopane)有抗病毒功效, 可作为治疗病毒感染的新型药物(Kumar and Siddique, Potential use of the Pteris vittata arsenic hyperaccumulation-regulation network for phytoremediation 1 2019 ... Yan等( Global transcriptome analysis of Huperzia serrata and identification of critical genes involved in the biosynthesis of huperzine A 1 2017 ... 生物碱是植物体内一类含氮杂环有机化合物.石松碱(即石松类植物生物碱的统称) (Siengalewicz et al., Metabolomic profiling in Selaginella lepidophylla at various hydration states provides new insights into the mechanistic basis of desiccation tolerance 1 2013 ... 石松类植物卷柏和一些蕨类具有极强的耐旱性(Alejo-Jacuind and Herrera-Estrella, Occurrence of brassinosteroids in non-flowering land plants, liverwort, moss, lycophyte and fern 2 2017 ... IAAs和细胞分裂素(CTKs)协同调控植物根系发育, 用外源IAA和CTK处理细叶满江红根系后, 根分生组织细胞数目分别减少和增多, 根细胞壁合成相关基因(CYP71A20、EXPANSIN、ABCB15、TOR1、IRX12和BAT1)的表达量发生改变, IAA (AfPINs、AfAUX、AfIAAs和AfARFs等)和CTK (AfAHK3、AfCRE1、AfAHPs和AfARRs)调动对应的激素信号通路发挥生物学功能(de Vries et al.,
- 未知 ... The ‘how’ and ‘where’ of plant microRNAs 1 2017 ... miRNAs是基因转录后关键调节因子, 主要通过mRNA剪切和翻译发挥作用, 在各类植物中普遍存在(Yu et al., Comparative analysis of arsenic transport and tolerance mechanisms: evolution from prokaryote to higher plants 1 2022 ... 基于已有的研究成果, Cai等( Isolation and functional characterization of two Caffeoyl Coenzyme A 3-O-methyltransferases from the fern species Polypodiodes amoena 1 2019 ... 体外酶活实验表明, 种子植物CYP98 (cytochrome P450 monooxygenase 98)的底物种类丰富, 4-香豆酰莽草酸酯是裸子植物火炬松(Pinus taeda) PtCYP98的最优催化底物; 但是江南卷柏和蜈蚣草CYP98s对底物4-香豆酰莽草酸酯催化效率低, 更易催化4-香豆酰邻氨基苯甲酸酯; 被子植物CYP98催化底物丰富多样, 为其适应复杂多变的生存环境提供了更多保护性物质(Alber et al., Genomic insights into genetic diploidization in the homosporous fern Adiantum nelumboides 1 2022 ... Pteridophytes with whole-genome sequence published
- 未知 ... Molecular cloning and characterization of three cryptochrome genes from the fern Asplenium yunnanense 1 2013 ... 植物通过隐花色素(CRY)感受蓝光并介导蓝光信号转导.CRY基因起源古老, 存在于从绿藻到种子植物的所有植物中(Cai et al., The evolution of the KANADI gene family and leaf development in lycophytes and ferns 1 2019 ... 蕨类植物是地球上较早出现的陆生维管植物, 是研究多成员家族基因起源与演化的关键植物类群; 绿色植物由简单到复杂的演化过程中, 不同物种基因组中家族基因的种类、数目和功能会发生改变.基因组测序显示美洲水蕨MADS-box基因家族有35个成员, 但蕨类植物演化至美洲水蕨这一分支出现时一些特异的MADS- box转录因子丢失(Marchant?? et al., /
本文的其它图/表
|
 首页
首页