植物性别决定基因及其表观遗传调控研究进展
罗兰莎, 宋雯佩, 化青珠, 李大卫, 梁红, 张宪智
植物学报
2024, 59 ( 2):
278-290.
DOI: 10.11983/CBB23088
雌雄异株植物的性别决定机制是繁殖生物学、进化与生态学等多个学科的前沿热点问题。近年来, 一些重要经济作物如芦笋(Asparagus officinalis)、猕猴桃(Actinidia spp.)和杨树(Populus spp.)的性别决定机制已被揭示, 并应用于性别特异性产品开发和两性新品种培育。该文先从植物性染色体和性别决定基因两方面系统分析植物性别决定的遗传学基础, 并深入讨论非编码RNA和DNA甲基化两类表观遗传调控途径在性别决定分子通路中的作用。在此基础上, 提出后续有待开展植物性别决定基因间的比较研究, 并深入解析植物性别决定中的表观遗传调控机制, 以深化对雌雄异株植物性别决定分子机制的认识并扩展其在农业生产上的应用价值。
| 分类群 | 物种 | 性别决定基因 | 参考文献 | | 猕猴桃科 | 中华猕猴桃(Actinidia chinensis) | SyGl: 雄性抑制基因
FrBy: 雄性激活基因 | Akagi et al., 2018, 2019 | | 番木瓜科 | 番木瓜(Carica papaya) | CpSVP: 功能未验证
CpSERK/CpCAF1AL: 功能未验证 | Urasaki et al., 2012 | | 百合科 | 石刁柏(Asparagus officinalis) | SOFF: 雌性抑制基因
AspTDF1: 雄性激活基因 | Harkess et al., 2017, 2020 | | 柿树科 | 君迁子(Diospyros lotus) | OGl: 雄性激活基因 | Akagi et al., 2014, 2020 | | 桑科 | 对叶榕(Ficus hispida) | FhAG2: 雄性激活基因 | Zhang et al., 2020 | | 蔷薇科 | 弗吉尼亚草莓(Fragaria virginiana) | GMEW: 功能未验证
RPP0W: 功能未验证 | Tennessen et al., 2018 | | 猪笼草科 | 小猪笼草(Nepenthes gracilis) | DYT1: 雄性激活基因
SEP1: 功能未验证 | Scharmann et al., 2019 | | 棕榈科 | 海枣(Phoenix dactylifera) | CYP703/GPAT3: 雄性激活基因
LOG-like: 雄性抑制基因 | Torres et al., 2018 | | 石竹科 | 白花蝇子草(Silene latifolia) | CLAVATA3-like: 雌性抑制基因 | Kazama et al., 2022 | | 葡萄科 | 葡萄(Vitis vinifera) | VvilNP1: 雄性激活基因
VviPLATZ1: 雌性激活基因 | Massonnet et al., 2020; Iocco- Corena et al., 2021 | | 苏铁科 | 攀枝花苏铁(Cycas panzhihuaensis) | CYCAS_034085: 功能未验证 | Liu et al., 2022 | | 银杏科 | 银杏(Ginkgo biloba) | GbMADS18: 功能未验证
Gb_15883/15884/15885/15886/
28587: 功能未验证 | Liao et al., 2020 | | 杨梅科 | 杨梅(Morella rubra) | MrCPS2: 功能未验证
MrASP2: 功能未验证 | Jia et al., 2019 | | 藜科 | 菠菜(Spinacia oleracea) | NRT1/PTR6.4: 雄性激活基因 | Ma et al., 2022 | | 杨柳科 | 银白杨(Populus alba)
欧洲山杨(P. tremula)
毛果杨(P. trichocarpa) | ARR17: 雄性抑制基因
ARR17-IR: 雌性抑制基因 | Müller et al., 2020 | | 美洲黑杨(P. deltoids) | FERR-IR: 雌性抑制基因
MSL: 雄性激活基因 | Xue et al., 2020 | | 胡杨(P. euphratica) | RR-IR: 雌性抑制基因 | Yang et al., 2021 | | 腺柳(Salix chaenomeloides)
钻天柳(S. arbutifolia)
三蕊柳(S. triandra) | RR: 雄性抑制基因 | Wang et al., 2022, 2023 | | 红皮柳(S. purpurea) | ARR17: 雄性抑制基因
GATA15: 雌性激活基因 | Hyden et al., 2023 |
View table in article
表2
雌雄异株植物的性别决定基因
正文中引用本图/表的段落
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础。从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2)。其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978)。在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花。Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy。SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别。Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1。Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性。Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性。在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021)。
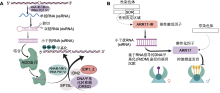
RNA指导的DNA甲基化(RNA-directed DNA methylation, RdDM)是植物特有的DNA从头甲基化途径, 在调控基因表达、沉默转座子和维持基因组稳定性方面发挥重要作用(邵凤清等, 2023)。该途径主要包括2个步骤: 首先, 在植物特有的RNA聚合酶IV (Pol IV)指导下转录产生小干扰RNA (siRNA)前体, 经由一系列辅助蛋白加工, 最终形成长度为24 nt的成熟siRNA; 其次, 24 nt siRNA与AGO4/6蛋白结合, 互补配对RNA聚合酶V (Pol V)转录形成的基因间非编码RNA (intergenic non-coding RNA, IGN RNA), 招募DNA甲基转移酶DRM2在相应位置从头建立DNA甲基化, 从而实现基因沉默(Erdmann and Picard, 2020)。在杨柳科植物中发现1个促进雌性、抑制雄性的基因ARR17 (Müller et al., 2020; Xue et al., 2020)。然而, 在一些杨属物种, 如欧洲山杨、毛果杨、美洲黑杨和胡杨(P. euphratica)中, 该基因并不位于Y染色体的性别决定区域(Y-SDR) (Yang et al., 2021)。相应地, 在Y-SDR区域存在ARR17基因的反向重复序列(ARR17-IR)。ARR17-IR可以转录产生长度为24 nt的siRNAs, 进一步通过RdDM途径引起ARR17启动子区域发生甲基化, 从而抑制ARR17的表达(图2)。在柳属中的腺柳和钻天柳中也发现Y- SDR存在ARR17-IR片段, 表明ARR17-IR通过RdDM途径调控ARR17基因表达的性别决定方式在杨柳科植物中具有一定的保守性(Wang et al., 2022)。
植物性别决定一直是遗传学、发育生物学、进化与生态学研究的重点方向。雌雄异株植物分布在植物生命之树的不同谱系中, 为多次独立发生, 是研究性染色体起源与演化的理想体系(Ming et al., 2011; 秦力等, 2016)。2014年克隆了第1个植物性别决定基因OGI, 至今已从猕猴桃、芦笋、葡萄、杨树和柳树等多个物种的性染色体中鉴定了性别决定基因(表1, 表2), 为不同植物类群间性别决定基因的比较研究奠定了基础。通过比较基因功能, 发现促雄基因(如芦笋的AspTDF1和猕猴桃的FrBy)均为花粉绒毡层发育关键基因; 类似地, 杨树ARR17和猕猴桃SyGl均涉及细胞分裂素信号通路(Leite Montalv?o et al., 2021)。不同植物的非同源性别决定基因参与相同的生物学通路, 表明植物的性别决定基因可能具有一定的功能趋同进化。
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Classification and comparison of small RNAs from plants 2 2013 ... 非编码RNA是一类不编码蛋白质的RNA, 依据其结构大小可分为小干扰RNA (small-interfering RNA, siRNA)、微小RNA (microRNA, miRNA)和长链非编码RNA (long non-coding RNA, lncRNA)等.SiRNA是由双链RNA (double-stranded RNA, dsRNA)前体经核酸内切酶DCL产生的小RNA, 长度一般为21-22 nt (Axtell, 2013).成熟的siRNA分子呈双链, 与AGO蛋白结合形成RNA诱导沉默复合体(RNA-induced silencing complex, RISC).再进一步与靶基因的mRNA互补配对, 导致mRNA降解或者抑制其翻译, 从而实现靶基因沉默, 称之为RNA干扰(RNA interference, RNAi) (Hung and Slotkin, 2021).柿属物种具有1个雌性化基因MeGI, 若MeGI正常表达, 单性花表现为雌性; 反之则为雄性(Akagi et al., 2014).柿属二倍体物种君迁子是雌雄异株植物, 其Y染色体的性别决定区域(Y-SDR)包含1个MeGI的同源基因OGI.该基因末端发生反向重复(inverted repeat, IR)导致假基因化而失去编码蛋白质的功能.然而, OGI两端的反向重复序列可以转录形成发夹结构的双链RNA, 生成siRNA后通过RNAi途径特异性地沉默其常染色体上的同源基因MeGI, 最终决定个体发育为雄性(图1). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Multiple developmental processes underlie sex differentiation in angiosperms 1 2011 ... 雌雄异株(dioecy)是一种单性花在个体上分离的性别系统.在被子植物中, 约有15 600种(占6%)雌雄异株植物, 隶属于175科987属(Hobza et al., 2018).单性花的形成涉及一系列复杂的过程, 依据其发育模式可分为2种类型: I型单性花(Type I)和II型单性花(Type II).其中I型单性花在发育初期兼具雌蕊原基和雄蕊原基, 但在发育中后期雌蕊或者雄蕊败育, 最终形成仅有1种功能性器官的单性花(Diggle et al., 2011).大多数雌雄异株植物属于I型单性花, 即雄花中存在退化的雌蕊、雌花中存在败育的雄蕊, 如猕猴桃(Actinidia spp.)和柿(Diospyros spp.).II型单性花则不经历两性阶段, 在花分生组织中仅形成雄蕊或者雌蕊原基, 成熟花中无相反性器官的残留, 如杨柳科(Salicaceae)和杜仲科(Eucommiaceae)植物(陆静等, 2021). ... RNA-directed DNA methylation 3 2020 ... RNA指导的DNA甲基化(RNA-directed DNA methylation, RdDM)是植物特有的DNA从头甲基化途径, 在调控基因表达、沉默转座子和维持基因组稳定性方面发挥重要作用(邵凤清等, 2023).该途径主要包括2个步骤: 首先, 在植物特有的RNA聚合酶IV (Pol IV)指导下转录产生小干扰RNA (siRNA)前体, 经由一系列辅助蛋白加工, 最终形成长度为24 nt的成熟siRNA; 其次, 24 nt siRNA与AGO4/6蛋白结合, 互补配对RNA聚合酶V (Pol V)转录形成的基因间非编码RNA (intergenic non-coding RNA, IGN RNA), 招募DNA甲基转移酶DRM2在相应位置从头建立DNA甲基化, 从而实现基因沉默(Erdmann and Picard, 2020).在杨柳科植物中发现1个促进雌性、抑制雄性的基因ARR17 (Müller et al., 2020; Xue et al., 2020).然而, 在一些杨属物种, 如欧洲山杨、毛果杨、美洲黑杨和胡杨(P. euphratica)中, 该基因并不位于Y染色体的性别决定区域(Y-SDR) (Yang et al., 2021).相应地, 在Y-SDR区域存在ARR17基因的反向重复序列(ARR17-IR).ARR17-IR可以转录产生长度为24 nt的siRNAs, 进一步通过RdDM途径引起ARR17启动子区域发生甲基化, 从而抑制ARR17的表达(图2).在柳属中的腺柳和钻天柳中也发现Y- SDR存在ARR17-IR片段, 表明ARR17-IR通过RdDM途径调控ARR17基因表达的性别决定方式在杨柳科植物中具有一定的保守性(Wang et al., 2022). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Research progress of plant sex differentiation 1 2022 ... 性染色体是雌雄异株性系统形成和维持的根本原因, 其起源于1对常染色体, 在进化过程中其功能逐渐特化为决定植物的性别(李书粉等, 2015).Ming等(2011)通过分析雌雄异株植物的细胞遗传和分子遗传学数据, 提出性染色体进化涉及性别决定基因的产生和性别决定区域(sex determination region, SDR)的形成, 其中性染色体之间(如X与Y)的重组抑制是性别决定区域形成的必要条件.重组抑制表现为同源染色体不能正常进行交叉重组, 是常染色体进化为性染色体的前提(傅靖棋等, 2022).染色体倒位和异染色质化是重组抑制发生的首要原因, 重复序列的积累进而加剧性染色体之间(如X与Y)的重组抑制, 最终导致性别决定区域的产生以及Y染色体的异型化(Rifkin et al., 2021). ... 植物性别分化研究进展 1 2022 ... 性染色体是雌雄异株性系统形成和维持的根本原因, 其起源于1对常染色体, 在进化过程中其功能逐渐特化为决定植物的性别(李书粉等, 2015).Ming等(2011)通过分析雌雄异株植物的细胞遗传和分子遗传学数据, 提出性染色体进化涉及性别决定基因的产生和性别决定区域(sex determination region, SDR)的形成, 其中性染色体之间(如X与Y)的重组抑制是性别决定区域形成的必要条件.重组抑制表现为同源染色体不能正常进行交叉重组, 是常染色体进化为性染色体的前提(傅靖棋等, 2022).染色体倒位和异染色质化是重组抑制发生的首要原因, 重复序列的积累进而加剧性染色体之间(如X与Y)的重组抑制, 最终导致性别决定区域的产生以及Y染色体的异型化(Rifkin et al., 2021). ... lncRNAs in plant and animal sexual reproduction 1 2018 ... LncRNA是长度超过200 nt的非编码RNA, 近年来的研究发现, lncRNA参与植物的性别决定(Golicz et al., 2018).在水稻(Oryza sativa)中, 部分lncRNA可充当miRNA的前体, 影响雌配子体发育和花粉育性(Liu et al., 2019).玉米(Zea mays) Zm401-lncRNA在花粉绒毡层中特异性表达, 其突变后导致雄性不育(Ma et al., 2008).在黄瓜(Cucumis sativus)中, CSM10-lncRNA促使雄花分化(Cho et al., 2005).近期, 对杜仲的花芽进行链特异性文库测序, 经比较分析发现雌、雄间差异表达的312个lncRNA.通过lncRNA的调控网络分析, 共鉴定到5个lncRNA (LNC_025233、LNC_017049、TUCP_009073、TUCP_009932和TUCP_001849)可能参与杜仲的性别决定(朱利利, 2019). ... Sex-determining chromosomes and sexual dimorphism: insights from genetic mapping of sex expression in a natural hybrid Fragaria × ananassa subsp. cuneifolia 1 2013 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Sex determination by two Y-linked genes in garden asparagus 2 2020 ... 植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
与猕猴桃、芦笋和葡萄的“双基因”性别决定机制不同, 柿属(Diospyros)、杨属(Populus)和柳属(Salix)物种的性别由性别决定区域的1个基因控制, 称之为“单基因模型”(Renner, 2016).Akagi等(2014)结合基因组和转录组测序, 从君迁子(D. lotus)中鉴定到1个位于Y-SDR区域的性别决定基因OGI.该基因通过抑制其常染色体上旁系同源基因MeGI的表达控制花的性别.Müller等(2020)通过基因组重测序及全基因组关联分析, 鉴定了欧洲山杨(P. tremula)、毛果杨(P. trichocarpa)和银白杨(P. alba)的1个雌性化基因ARR17, 该基因起促雌和抑雄双重作用.利用CRISPR/Cas技术敲除ARR17导致雌株发生性别转变而开出雄花.Xue等(2020)对美洲黑杨(P. deltoides)的研究进一步证实ARR17 (FERR)促进雌蕊发育.在腺柳(S. chaenomeloides)、钻天柳(S. arbutifolia)、三蕊柳(S. triandra)和红皮柳(S. purpurea)的性别决定区域也发现了ARR17的同源基因, 表明该基因在杨柳科植物中的功能保守(Wang et al., 2022, 2023; Hyden et al., 2023). ... The asparagus genome sheds light on the origin and evolution of a young Y chromosome 2 2017 ... 植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Genetic degeneration of old and young Y chromosomes in the flowering plant Rumex hastatulus 1 2014 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... The initiation of RNA interference (RNAi) in plants 3 2021 ... 非编码RNA是一类不编码蛋白质的RNA, 依据其结构大小可分为小干扰RNA (small-interfering RNA, siRNA)、微小RNA (microRNA, miRNA)和长链非编码RNA (long non-coding RNA, lncRNA)等.SiRNA是由双链RNA (double-stranded RNA, dsRNA)前体经核酸内切酶DCL产生的小RNA, 长度一般为21-22 nt (Axtell, 2013).成熟的siRNA分子呈双链, 与AGO蛋白结合形成RNA诱导沉默复合体(RNA-induced silencing complex, RISC).再进一步与靶基因的mRNA互补配对, 导致mRNA降解或者抑制其翻译, 从而实现靶基因沉默, 称之为RNA干扰(RNA interference, RNAi) (Hung and Slotkin, 2021).柿属物种具有1个雌性化基因MeGI, 若MeGI正常表达, 单性花表现为雌性; 反之则为雄性(Akagi et al., 2014).柿属二倍体物种君迁子是雌雄异株植物, 其Y染色体的性别决定区域(Y-SDR)包含1个MeGI的同源基因OGI.该基因末端发生反向重复(inverted repeat, IR)导致假基因化而失去编码蛋白质的功能.然而, OGI两端的反向重复序列可以转录形成发夹结构的双链RNA, 生成siRNA后通过RNAi途径特异性地沉默其常染色体上的同源基因MeGI, 最终决定个体发育为雄性(图1). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Molecular cytogenetics in hop (Humulus lupulus L.) and identification of sex chromosomes by DAPI-banding 1 2003 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... A CLAVATA3-like gene acts as a gynoecium suppression function in white campion 1 2022 ... Sex determination genes of dioecious plants
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... The quest for epigenetic regulation underlying unisexual flower development in Cucumis melo 1 2017 ... DNA甲基化是一种常见的植物表观遗传修饰, 发生在对称的CG和CHG, 以及非对称的CHH上(H=A/T/ C), 调控基因的表达(邵凤清等, 2023).前期研究已发现DNA甲基化影响植物开花过程以及花的性别.例如, 拟南芥(Arabidopsis thaliana)的FWA编码开花抑制因子, 当FWA上游重复序列被甲基化修饰时, FWA基因受到抑制, 植株开花时间提前(齐鲁琦, 2018).Latrasse等(2017)在甜瓜(C. melo)中发现1个转座子元件的插入, 使得CmWIP1启动子发生DNA甲基化, 从而导致雄花转变为雌花.低温处理黄瓜茎尖会引起基因组DNA甲基化水平下降, 导致黄瓜的雌花数量明显增多(周赓等, 2019).Akagi等(2016)对雌雄异花同株(monoecy)的六倍体栽培柿(D. kaki)展开研究, 结果表明栽培柿雄花MeGI甲基化程度显著高于雌花, 抑制了其在雄花中的表达从而使雄蕊正常发育.同时OGI启动子区域有一段268 bp的转座子元件插入, 导致该基因由于发生较高程度的甲基化而抑制其表达, 进而产生雌花.与二倍体君迁子相比, 六倍体栽培柿通过DNA甲基化更灵活地调控单性花的发育. ... The diversity and dynamics of sex determination in dioecious plants 1 2021 ... 植物性别决定一直是遗传学、发育生物学、进化与生态学研究的重点方向.雌雄异株植物分布在植物生命之树的不同谱系中, 为多次独立发生, 是研究性染色体起源与演化的理想体系(Ming et al., 2011; 秦力等, 2016).2014年克隆了第1个植物性别决定基因OGI, 至今已从猕猴桃、芦笋、葡萄、杨树和柳树等多个物种的性染色体中鉴定了性别决定基因(表1, 表2), 为不同植物类群间性别决定基因的比较研究奠定了基础.通过比较基因功能, 发现促雄基因(如芦笋的AspTDF1和猕猴桃的FrBy)均为花粉绒毡层发育关键基因; 类似地, 杨树ARR17和猕猴桃SyGl均涉及细胞分裂素信号通路(Leite Montalv?o et al., 2021).不同植物的非同源性别决定基因参与相同的生物学通路, 表明植物的性别决定基因可能具有一定的功能趋同进化. ... ARR17 controls dioecy in Populus by repressing B-class MADS-box gene expression 1 2022 ... 此外, 多个不同植物类群的性别决定研究结果也为雌雄异株植物性别决定分子通路的进化分析提供了基础数据.通过比较杨树和柿等不同类群的性别决定机制, 发现性别决定基因与花发育MADS-box基因之间存在直接或间接的调控关系, 表明不同类群之间很可能共用一条保守的性别调控路径(Yang et al., 2019; Leite Montalv?o et al., 2022).在该性别决定的保守模型中, 植物的性别决定基因位于性染色体的性别决定区域(Y-SDR)中, 通过调控下游基因(如杨树的ARR17和UFO及君迁子的MeGI和SVP)的表达, 最终影响花发育ABC模型中B类基因(AP3或PI)的表达, 决定单性花的性别(Zhang et al., 2022). ... 1 2014 ... MiRNA是一类长度为20-24 nt的非编码RNA, 可通过碱基互补配对的方式与靶基因mRNA结合, 使靶基因mRNA降解或者抑制其翻译, 实现miRNA对靶基因的负调控(Axtell, 2013).前期研究发现, miR172和miR169可控制花发育ABC类基因的表达, 进而控制矮牵牛(Petunia hybrida)和金鱼草(Antirrhinum majus)的花器官表型(Cartolano et al., 2007).近期, 有关雌雄异株植物的研究也发现miRNA在雌、雄个体间差异表达.例如, 在番木瓜花中共检测到65个miRNA, 其中14个miRNA在雌、雄花间差异表达(李崇奇, 2014).在油棕(Elaeis guineensis)中, miRNA159a表现出雌性偏向表达式样, 可能参与雌蕊发育(Somyong et al., 2016).对中华猕猴桃雌、雄花的小RNA组学分析显示, 170个miRNA在性别间差异表达, 并且在猕猴桃的性染色体(即Chr. 25)上预测到3个miRNA, 其中novel-ach-miR362的靶基因Achn298021与猕猴桃的单性花发育有关(闫明科等, 2015).此外, 对杜仲(Eucommia ulmoides)花芽进行小RNA测序, 共鉴定到187个miRNA, 其中31个为性别间差异表达的miRNA.进一步的靶基因预测分析显示, novel-23-miRNA调控花发育ABC类基因DEF的表达, 可能参与杜仲的性别决定(朱利利, 2019). ... 1 2014 ... MiRNA是一类长度为20-24 nt的非编码RNA, 可通过碱基互补配对的方式与靶基因mRNA结合, 使靶基因mRNA降解或者抑制其翻译, 实现miRNA对靶基因的负调控(Axtell, 2013).前期研究发现, miR172和miR169可控制花发育ABC类基因的表达, 进而控制矮牵牛(Petunia hybrida)和金鱼草(Antirrhinum majus)的花器官表型(Cartolano et al., 2007).近期, 有关雌雄异株植物的研究也发现miRNA在雌、雄个体间差异表达.例如, 在番木瓜花中共检测到65个miRNA, 其中14个miRNA在雌、雄花间差异表达(李崇奇, 2014).在油棕(Elaeis guineensis)中, miRNA159a表现出雌性偏向表达式样, 可能参与雌蕊发育(Somyong et al., 2016).对中华猕猴桃雌、雄花的小RNA组学分析显示, 170个miRNA在性别间差异表达, 并且在猕猴桃的性染色体(即Chr. 25)上预测到3个miRNA, 其中novel-ach-miR362的靶基因Achn298021与猕猴桃的单性花发育有关(闫明科等, 2015).此外, 对杜仲(Eucommia ulmoides)花芽进行小RNA测序, 共鉴定到187个miRNA, 其中31个为性别间差异表达的miRNA.进一步的靶基因预测分析显示, novel-23-miRNA调控花发育ABC类基因DEF的表达, 可能参与杜仲的性别决定(朱利利, 2019). ... A rapid method for assembly of single chromosome and identification of sex determination region based on single-chromosome sequencing 2 2023a ... 大多数植物的性染色体处于早期演化阶段, 无法从形态上直接区分性染色体和常染色体, 且同源性染色体之间(如X与Y)也无明显的形态差别, 通过核型分析难以确定(Baránková et al., 2020).此外, 由于性染色体上的性别决定区域缺乏遗传重组, 基于传统的重组定位方法难以获得该区域的具体遗传信息(彭丹和武志强, 2022).伴随着基因组测序和生物信息学分析方法的发展, 近年来逐步解析了一些雌雄异株植物的基因组, 并基于基因组数据鉴定了性染色体组成及其性别决定区域.据不完全统计, 截至目前已完成基因组测序的雌雄异株植物共计35种, 其中22种具有XX/XY性染色体, 10种具有ZW/ZZ性染色体, 另外3种苔藓植物具有U/V性染色体(表1).然而, 目前多数雌雄异株植物的基因组学研究仅选取1个雌性或者雄性个体进行测序, 获得的基因组数据中遗失性染色体信息; 且由于测序技术所限, 无法跨越性染色体的大片段重复序列区域, 难以实现性染色体(如X与Y)的分型组装.近年来, 基于长读长的测序技术和分析方法, 获得了高度连续的人类Y染色体序列和端粒到端粒(T2T)的完整X染色体序列(Miga et al., 2020).Li等(2023a)通过分离菠菜的X与Y染色体, 并对单条染色体进行测序, 分别获得192.1 Mb的X染色体序列和195.2 Mb的Y染色体序列.上述研究为植物性染色体的测序与分析提供了技术参考.在T2T基因组时代, 实现雄性异配个体X与Y或者雌性异配个体Z与W的染色体分型组装是未来的重点研究方向.高质量的单倍型参考基因组也是进行性别决定基因鉴定和探讨植物性二型差异的基础. ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Role of transposons in origin and evolution of plant XY sex chromosomes 1 2015 ... 性染色体是雌雄异株性系统形成和维持的根本原因, 其起源于1对常染色体, 在进化过程中其功能逐渐特化为决定植物的性别(李书粉等, 2015).Ming等(2011)通过分析雌雄异株植物的细胞遗传和分子遗传学数据, 提出性染色体进化涉及性别决定基因的产生和性别决定区域(sex determination region, SDR)的形成, 其中性染色体之间(如X与Y)的重组抑制是性别决定区域形成的必要条件.重组抑制表现为同源染色体不能正常进行交叉重组, 是常染色体进化为性染色体的前提(傅靖棋等, 2022).染色体倒位和异染色质化是重组抑制发生的首要原因, 重复序列的积累进而加剧性染色体之间(如X与Y)的重组抑制, 最终导致性别决定区域的产生以及Y染色体的异型化(Rifkin et al., 2021). ... 转座子在植物XY性染色体起源与演化过程中的作用 1 2015 ... 性染色体是雌雄异株性系统形成和维持的根本原因, 其起源于1对常染色体, 在进化过程中其功能逐渐特化为决定植物的性别(李书粉等, 2015).Ming等(2011)通过分析雌雄异株植物的细胞遗传和分子遗传学数据, 提出性染色体进化涉及性别决定基因的产生和性别决定区域(sex determination region, SDR)的形成, 其中性染色体之间(如X与Y)的重组抑制是性别决定区域形成的必要条件.重组抑制表现为同源染色体不能正常进行交叉重组, 是常染色体进化为性染色体的前提(傅靖棋等, 2022).染色体倒位和异染色质化是重组抑制发生的首要原因, 重复序列的积累进而加剧性染色体之间(如X与Y)的重组抑制, 最终导致性别决定区域的产生以及Y染色体的异型化(Rifkin et al., 2021). ... DNA methylation is involved in sexual differentiation and sex chromosome evolution in the dioecious plant garden asparagus 2 2021 ... 雌雄异株植物的性别决定机制一直是生物学领域的研究热点, 对于理解性染色体的起源与演化及培育两性新品种具有重要意义(Zhang et al., 2022).对于以获取果实和种子为目的的植物, 生产上偏向利用雌株, 如猕猴桃、开心果(即阿月浑子(Pistacia vera))、杨梅(Morella rubra)、柿和番木瓜(Carica papaya); 而对于以收获茎叶为目的的芦笋(即石刁柏(Asparagus officinalis))和菠菜(Spinacia oleracea)等蔬菜来说, 则主要种植雄株(赵玉洁等, 2018).近年来, 随着基因组学的发展以及CRISPR/Cas基因编辑技术的应用, 越来越多的性染色体及位于其上的性别决定基因得以揭示, 阐明了雌雄异株植物性别决定的分子基础.特别是最新研究揭示了表观遗传调控(如非编码RNA和DNA甲基化)在植物性别决定通路中发挥重要作用(Akagi et al., 2020; Xue et al., 2020; Li et al., 2021).本文详细梳理了近年来雌雄异株植物的性染色体与性别决定基因研究成果, 分析了不同植物性别决定的遗传学基础; 并进一步从表观遗传学角度阐述了非编码RNA和DNA甲基化在植物性别决定中的调控作用, 以期为深入理解雌雄异株植物的起源与演化及保护利用提供参考依据. ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... The genomic architecture of the sex-determining region and sex-related metabolic variation in Ginkgo biloba 2 2020 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Research progress on sex determination genes of woody plants 1 2021 ... 雌雄异株(dioecy)是一种单性花在个体上分离的性别系统.在被子植物中, 约有15 600种(占6%)雌雄异株植物, 隶属于175科987属(Hobza et al., 2018).单性花的形成涉及一系列复杂的过程, 依据其发育模式可分为2种类型: I型单性花(Type I)和II型单性花(Type II).其中I型单性花在发育初期兼具雌蕊原基和雄蕊原基, 但在发育中后期雌蕊或者雄蕊败育, 最终形成仅有1种功能性器官的单性花(Diggle et al., 2011).大多数雌雄异株植物属于I型单性花, 即雄花中存在退化的雌蕊、雌花中存在败育的雄蕊, 如猕猴桃(Actinidia spp.)和柿(Diospyros spp.).II型单性花则不经历两性阶段, 在花分生组织中仅形成雄蕊或者雌蕊原基, 成熟花中无相反性器官的残留, 如杨柳科(Salicaceae)和杜仲科(Eucommiaceae)植物(陆静等, 2021). ... 木本植物性别决定基因研究进展 1 2021 ... 雌雄异株(dioecy)是一种单性花在个体上分离的性别系统.在被子植物中, 约有15 600种(占6%)雌雄异株植物, 隶属于175科987属(Hobza et al., 2018).单性花的形成涉及一系列复杂的过程, 依据其发育模式可分为2种类型: I型单性花(Type I)和II型单性花(Type II).其中I型单性花在发育初期兼具雌蕊原基和雄蕊原基, 但在发育中后期雌蕊或者雄蕊败育, 最终形成仅有1种功能性器官的单性花(Diggle et al., 2011).大多数雌雄异株植物属于I型单性花, 即雄花中存在退化的雌蕊、雌花中存在败育的雄蕊, 如猕猴桃(Actinidia spp.)和柿(Diospyros spp.).II型单性花则不经历两性阶段, 在花分生组织中仅形成雄蕊或者雌蕊原基, 成熟花中无相反性器官的残留, 如杨柳科(Salicaceae)和杜仲科(Eucommiaceae)植物(陆静等, 2021). ... Zm401, a short-open reading-frame mRNA or noncoding RNA, is essential for tapetum and microspore development and can regulate the floret formation in maize 1 2008 ... LncRNA是长度超过200 nt的非编码RNA, 近年来的研究发现, lncRNA参与植物的性别决定(Golicz et al., 2018).在水稻(Oryza sativa)中, 部分lncRNA可充当miRNA的前体, 影响雌配子体发育和花粉育性(Liu et al., 2019).玉米(Zea mays) Zm401-lncRNA在花粉绒毡层中特异性表达, 其突变后导致雄性不育(Ma et al., 2008).在黄瓜(Cucumis sativus)中, CSM10-lncRNA促使雄花分化(Cho et al., 2005).近期, 对杜仲的花芽进行链特异性文库测序, 经比较分析发现雌、雄间差异表达的312个lncRNA.通过lncRNA的调控网络分析, 共鉴定到5个lncRNA (LNC_025233、LNC_017049、TUCP_009073、TUCP_009932和TUCP_001849)可能参与杜仲的性别决定(朱利利, 2019). ... The spinach YY genome reveals sex chromosome evolution, domestication, and introgression history of the species 1 2022 ... Sex determination genes of dioecious plants
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... The genetic basis of sex determination in grapes 2 2020 ... 植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Telomere-to-te- lomere assembly of a complete human X chromosome 1 2020 ... 大多数植物的性染色体处于早期演化阶段, 无法从形态上直接区分性染色体和常染色体, 且同源性染色体之间(如X与Y)也无明显的形态差别, 通过核型分析难以确定(Baránková et al., 2020).此外, 由于性染色体上的性别决定区域缺乏遗传重组, 基于传统的重组定位方法难以获得该区域的具体遗传信息(彭丹和武志强, 2022).伴随着基因组测序和生物信息学分析方法的发展, 近年来逐步解析了一些雌雄异株植物的基因组, 并基于基因组数据鉴定了性染色体组成及其性别决定区域.据不完全统计, 截至目前已完成基因组测序的雌雄异株植物共计35种, 其中22种具有XX/XY性染色体, 10种具有ZW/ZZ性染色体, 另外3种苔藓植物具有U/V性染色体(表1).然而, 目前多数雌雄异株植物的基因组学研究仅选取1个雌性或者雄性个体进行测序, 获得的基因组数据中遗失性染色体信息; 且由于测序技术所限, 无法跨越性染色体的大片段重复序列区域, 难以实现性染色体(如X与Y)的分型组装.近年来, 基于长读长的测序技术和分析方法, 获得了高度连续的人类Y染色体序列和端粒到端粒(T2T)的完整X染色体序列(Miga et al., 2020).Li等(2023a)通过分离菠菜的X与Y染色体, 并对单条染色体进行测序, 分别获得192.1 Mb的X染色体序列和195.2 Mb的Y染色体序列.上述研究为植物性染色体的测序与分析提供了技术参考.在T2T基因组时代, 实现雄性异配个体X与Y或者雌性异配个体Z与W的染色体分型组装是未来的重点研究方向.高质量的单倍型参考基因组也是进行性别决定基因鉴定和探讨植物性二型差异的基础. ... Sex chromosomes in land plants 2 2011 ... 性染色体是雌雄异株性系统形成和维持的根本原因, 其起源于1对常染色体, 在进化过程中其功能逐渐特化为决定植物的性别(李书粉等, 2015).Ming等(2011)通过分析雌雄异株植物的细胞遗传和分子遗传学数据, 提出性染色体进化涉及性别决定基因的产生和性别决定区域(sex determination region, SDR)的形成, 其中性染色体之间(如X与Y)的重组抑制是性别决定区域形成的必要条件.重组抑制表现为同源染色体不能正常进行交叉重组, 是常染色体进化为性染色体的前提(傅靖棋等, 2022).染色体倒位和异染色质化是重组抑制发生的首要原因, 重复序列的积累进而加剧性染色体之间(如X与Y)的重组抑制, 最终导致性别决定区域的产生以及Y染色体的异型化(Rifkin et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... A single gene underlies the dynamic evolution of poplar sex determination 6 2020 ... Sex determination genes of dioecious plants
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Genetic mapping of linkage group XIX and identification of sex-linked SSR markers in a Populus tremula × Populus tremuloides cross 1 2011 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Genetic linkage maps of Populus alba L. and comparative mapping analysis of sex determination across Populus species 1 2010 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Progress on sex determination of dioecious plants 1 2022 ... 大多数植物的性染色体处于早期演化阶段, 无法从形态上直接区分性染色体和常染色体, 且同源性染色体之间(如X与Y)也无明显的形态差别, 通过核型分析难以确定(Baránková et al., 2020).此外, 由于性染色体上的性别决定区域缺乏遗传重组, 基于传统的重组定位方法难以获得该区域的具体遗传信息(彭丹和武志强, 2022).伴随着基因组测序和生物信息学分析方法的发展, 近年来逐步解析了一些雌雄异株植物的基因组, 并基于基因组数据鉴定了性染色体组成及其性别决定区域.据不完全统计, 截至目前已完成基因组测序的雌雄异株植物共计35种, 其中22种具有XX/XY性染色体, 10种具有ZW/ZZ性染色体, 另外3种苔藓植物具有U/V性染色体(表1).然而, 目前多数雌雄异株植物的基因组学研究仅选取1个雌性或者雄性个体进行测序, 获得的基因组数据中遗失性染色体信息; 且由于测序技术所限, 无法跨越性染色体的大片段重复序列区域, 难以实现性染色体(如X与Y)的分型组装.近年来, 基于长读长的测序技术和分析方法, 获得了高度连续的人类Y染色体序列和端粒到端粒(T2T)的完整X染色体序列(Miga et al., 2020).Li等(2023a)通过分离菠菜的X与Y染色体, 并对单条染色体进行测序, 分别获得192.1 Mb的X染色体序列和195.2 Mb的Y染色体序列.上述研究为植物性染色体的测序与分析提供了技术参考.在T2T基因组时代, 实现雄性异配个体X与Y或者雌性异配个体Z与W的染色体分型组装是未来的重点研究方向.高质量的单倍型参考基因组也是进行性别决定基因鉴定和探讨植物性二型差异的基础. ... 植物雌雄异株性别决定研究进展 1 2022 ... 大多数植物的性染色体处于早期演化阶段, 无法从形态上直接区分性染色体和常染色体, 且同源性染色体之间(如X与Y)也无明显的形态差别, 通过核型分析难以确定(Baránková et al., 2020).此外, 由于性染色体上的性别决定区域缺乏遗传重组, 基于传统的重组定位方法难以获得该区域的具体遗传信息(彭丹和武志强, 2022).伴随着基因组测序和生物信息学分析方法的发展, 近年来逐步解析了一些雌雄异株植物的基因组, 并基于基因组数据鉴定了性染色体组成及其性别决定区域.据不完全统计, 截至目前已完成基因组测序的雌雄异株植物共计35种, 其中22种具有XX/XY性染色体, 10种具有ZW/ZZ性染色体, 另外3种苔藓植物具有U/V性染色体(表1).然而, 目前多数雌雄异株植物的基因组学研究仅选取1个雌性或者雄性个体进行测序, 获得的基因组数据中遗失性染色体信息; 且由于测序技术所限, 无法跨越性染色体的大片段重复序列区域, 难以实现性染色体(如X与Y)的分型组装.近年来, 基于长读长的测序技术和分析方法, 获得了高度连续的人类Y染色体序列和端粒到端粒(T2T)的完整X染色体序列(Miga et al., 2020).Li等(2023a)通过分离菠菜的X与Y染色体, 并对单条染色体进行测序, 分别获得192.1 Mb的X染色体序列和195.2 Mb的Y染色体序列.上述研究为植物性染色体的测序与分析提供了技术参考.在T2T基因组时代, 实现雄性异配个体X与Y或者雌性异配个体Z与W的染色体分型组装是未来的重点研究方向.高质量的单倍型参考基因组也是进行性别决定基因鉴定和探讨植物性二型差异的基础. ... An efficient RNA-seq-based segregation analysis identifies the sex chromosomes of Cannabis sativa 1 2020 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Satellite DNA and transposable elements in seabuckthorn (Hippophae rhamnoides), a dioecious plant with small Y and large X chromosomes 1 2017 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... 1 2018 ... DNA甲基化是一种常见的植物表观遗传修饰, 发生在对称的CG和CHG, 以及非对称的CHH上(H=A/T/ C), 调控基因的表达(邵凤清等, 2023).前期研究已发现DNA甲基化影响植物开花过程以及花的性别.例如, 拟南芥(Arabidopsis thaliana)的FWA编码开花抑制因子, 当FWA上游重复序列被甲基化修饰时, FWA基因受到抑制, 植株开花时间提前(齐鲁琦, 2018).Latrasse等(2017)在甜瓜(C. melo)中发现1个转座子元件的插入, 使得CmWIP1启动子发生DNA甲基化, 从而导致雄花转变为雌花.低温处理黄瓜茎尖会引起基因组DNA甲基化水平下降, 导致黄瓜的雌花数量明显增多(周赓等, 2019).Akagi等(2016)对雌雄异花同株(monoecy)的六倍体栽培柿(D. kaki)展开研究, 结果表明栽培柿雄花MeGI甲基化程度显著高于雌花, 抑制了其在雄花中的表达从而使雄蕊正常发育.同时OGI启动子区域有一段268 bp的转座子元件插入, 导致该基因由于发生较高程度的甲基化而抑制其表达, 进而产生雌花.与二倍体君迁子相比, 六倍体栽培柿通过DNA甲基化更灵活地调控单性花的发育. ... 1 2018 ... DNA甲基化是一种常见的植物表观遗传修饰, 发生在对称的CG和CHG, 以及非对称的CHH上(H=A/T/ C), 调控基因的表达(邵凤清等, 2023).前期研究已发现DNA甲基化影响植物开花过程以及花的性别.例如, 拟南芥(Arabidopsis thaliana)的FWA编码开花抑制因子, 当FWA上游重复序列被甲基化修饰时, FWA基因受到抑制, 植株开花时间提前(齐鲁琦, 2018).Latrasse等(2017)在甜瓜(C. melo)中发现1个转座子元件的插入, 使得CmWIP1启动子发生DNA甲基化, 从而导致雄花转变为雌花.低温处理黄瓜茎尖会引起基因组DNA甲基化水平下降, 导致黄瓜的雌花数量明显增多(周赓等, 2019).Akagi等(2016)对雌雄异花同株(monoecy)的六倍体栽培柿(D. kaki)展开研究, 结果表明栽培柿雄花MeGI甲基化程度显著高于雌花, 抑制了其在雄花中的表达从而使雄蕊正常发育.同时OGI启动子区域有一段268 bp的转座子元件插入, 导致该基因由于发生较高程度的甲基化而抑制其表达, 进而产生雌花.与二倍体君迁子相比, 六倍体栽培柿通过DNA甲基化更灵活地调控单性花的发育. ... Research progress in plant sex chromosome evolution and sex determination genes 1 2016 ... 植物性别决定一直是遗传学、发育生物学、进化与生态学研究的重点方向.雌雄异株植物分布在植物生命之树的不同谱系中, 为多次独立发生, 是研究性染色体起源与演化的理想体系(Ming et al., 2011; 秦力等, 2016).2014年克隆了第1个植物性别决定基因OGI, 至今已从猕猴桃、芦笋、葡萄、杨树和柳树等多个物种的性染色体中鉴定了性别决定基因(表1, 表2), 为不同植物类群间性别决定基因的比较研究奠定了基础.通过比较基因功能, 发现促雄基因(如芦笋的AspTDF1和猕猴桃的FrBy)均为花粉绒毡层发育关键基因; 类似地, 杨树ARR17和猕猴桃SyGl均涉及细胞分裂素信号通路(Leite Montalv?o et al., 2021).不同植物的非同源性别决定基因参与相同的生物学通路, 表明植物的性别决定基因可能具有一定的功能趋同进化. ... 植物性染色体进化及性别决定基因研究进展 1 2016 ... 植物性别决定一直是遗传学、发育生物学、进化与生态学研究的重点方向.雌雄异株植物分布在植物生命之树的不同谱系中, 为多次独立发生, 是研究性染色体起源与演化的理想体系(Ming et al., 2011; 秦力等, 2016).2014年克隆了第1个植物性别决定基因OGI, 至今已从猕猴桃、芦笋、葡萄、杨树和柳树等多个物种的性染色体中鉴定了性别决定基因(表1, 表2), 为不同植物类群间性别决定基因的比较研究奠定了基础.通过比较基因功能, 发现促雄基因(如芦笋的AspTDF1和猕猴桃的FrBy)均为花粉绒毡层发育关键基因; 类似地, 杨树ARR17和猕猴桃SyGl均涉及细胞分裂素信号通路(Leite Montalv?o et al., 2021).不同植物的非同源性别决定基因参与相同的生物学通路, 表明植物的性别决定基因可能具有一定的功能趋同进化. ... Pathways for making unisexual flowers and unisexual plants: moving beyond the “two mutations linked on one chromosome” model 1 2016 ... 与猕猴桃、芦笋和葡萄的“双基因”性别决定机制不同, 柿属(Diospyros)、杨属(Populus)和柳属(Salix)物种的性别由性别决定区域的1个基因控制, 称之为“单基因模型”(Renner, 2016).Akagi等(2014)结合基因组和转录组测序, 从君迁子(D. lotus)中鉴定到1个位于Y-SDR区域的性别决定基因OGI.该基因通过抑制其常染色体上旁系同源基因MeGI的表达控制花的性别.Müller等(2020)通过基因组重测序及全基因组关联分析, 鉴定了欧洲山杨(P. tremula)、毛果杨(P. trichocarpa)和银白杨(P. alba)的1个雌性化基因ARR17, 该基因起促雌和抑雄双重作用.利用CRISPR/Cas技术敲除ARR17导致雌株发生性别转变而开出雄花.Xue等(2020)对美洲黑杨(P. deltoides)的研究进一步证实ARR17 (FERR)促进雌蕊发育.在腺柳(S. chaenomeloides)、钻天柳(S. arbutifolia)、三蕊柳(S. triandra)和红皮柳(S. purpurea)的性别决定区域也发现了ARR17的同源基因, 表明该基因在杨柳科植物中的功能保守(Wang et al., 2022, 2023; Hyden et al., 2023). ... Widespread recombination suppression facilitates plant sex chromosome evolution 1 2021 ... 性染色体是雌雄异株性系统形成和维持的根本原因, 其起源于1对常染色体, 在进化过程中其功能逐渐特化为决定植物的性别(李书粉等, 2015).Ming等(2011)通过分析雌雄异株植物的细胞遗传和分子遗传学数据, 提出性染色体进化涉及性别决定基因的产生和性别决定区域(sex determination region, SDR)的形成, 其中性染色体之间(如X与Y)的重组抑制是性别决定区域形成的必要条件.重组抑制表现为同源染色体不能正常进行交叉重组, 是常染色体进化为性染色体的前提(傅靖棋等, 2022).染色体倒位和异染色质化是重组抑制发生的首要原因, 重复序列的积累进而加剧性染色体之间(如X与Y)的重组抑制, 最终导致性别决定区域的产生以及Y染色体的异型化(Rifkin et al., 2021). ... Sex is determined by XY chromosomes across the radiation of dioecious Nepenthes pitcher plants 1 2019 ... Sex determination genes of dioecious plants
与猕猴桃、芦笋和葡萄的“双基因”性别决定机制不同, 柿属(Diospyros)、杨属(Populus)和柳属(Salix)物种的性别由性别决定区域的1个基因控制, 称之为“单基因模型”(Renner, 2016).Akagi等(2014)结合基因组和转录组测序, 从君迁子(D. lotus)中鉴定到1个位于Y-SDR区域的性别决定基因OGI.该基因通过抑制其常染色体上旁系同源基因MeGI的表达控制花的性别.Müller等(2020)通过基因组重测序及全基因组关联分析, 鉴定了欧洲山杨(P. tremula)、毛果杨(P. trichocarpa)和银白杨(P. alba)的1个雌性化基因ARR17, 该基因起促雌和抑雄双重作用.利用CRISPR/Cas技术敲除ARR17导致雌株发生性别转变而开出雄花.Xue等(2020)对美洲黑杨(P. deltoides)的研究进一步证实ARR17 (FERR)促进雌蕊发育.在腺柳(S. chaenomeloides)、钻天柳(S. arbutifolia)、三蕊柳(S. triandra)和红皮柳(S. purpurea)的性别决定区域也发现了ARR17的同源基因, 表明该基因在杨柳科植物中的功能保守(Wang et al., 2022, 2023; Hyden et al., 2023). ... Advances in research of DNA methylation regulation during fruit ripening 1 2023 ... RNA指导的DNA甲基化(RNA-directed DNA methylation, RdDM)是植物特有的DNA从头甲基化途径, 在调控基因表达、沉默转座子和维持基因组稳定性方面发挥重要作用(邵凤清等, 2023).该途径主要包括2个步骤: 首先, 在植物特有的RNA聚合酶IV (Pol IV)指导下转录产生小干扰RNA (siRNA)前体, 经由一系列辅助蛋白加工, 最终形成长度为24 nt的成熟siRNA; 其次, 24 nt siRNA与AGO4/6蛋白结合, 互补配对RNA聚合酶V (Pol V)转录形成的基因间非编码RNA (intergenic non-coding RNA, IGN RNA), 招募DNA甲基转移酶DRM2在相应位置从头建立DNA甲基化, 从而实现基因沉默(Erdmann and Picard, 2020).在杨柳科植物中发现1个促进雌性、抑制雄性的基因ARR17 (Müller et al., 2020; Xue et al., 2020).然而, 在一些杨属物种, 如欧洲山杨、毛果杨、美洲黑杨和胡杨(P. euphratica)中, 该基因并不位于Y染色体的性别决定区域(Y-SDR) (Yang et al., 2021).相应地, 在Y-SDR区域存在ARR17基因的反向重复序列(ARR17-IR).ARR17-IR可以转录产生长度为24 nt的siRNAs, 进一步通过RdDM途径引起ARR17启动子区域发生甲基化, 从而抑制ARR17的表达(图2).在柳属中的腺柳和钻天柳中也发现Y- SDR存在ARR17-IR片段, 表明ARR17-IR通过RdDM途径调控ARR17基因表达的性别决定方式在杨柳科植物中具有一定的保守性(Wang et al., 2022). ... 果实成熟过程中的DNA甲基化调控研究进展 1 2023 ... RNA指导的DNA甲基化(RNA-directed DNA methylation, RdDM)是植物特有的DNA从头甲基化途径, 在调控基因表达、沉默转座子和维持基因组稳定性方面发挥重要作用(邵凤清等, 2023).该途径主要包括2个步骤: 首先, 在植物特有的RNA聚合酶IV (Pol IV)指导下转录产生小干扰RNA (siRNA)前体, 经由一系列辅助蛋白加工, 最终形成长度为24 nt的成熟siRNA; 其次, 24 nt siRNA与AGO4/6蛋白结合, 互补配对RNA聚合酶V (Pol V)转录形成的基因间非编码RNA (intergenic non-coding RNA, IGN RNA), 招募DNA甲基转移酶DRM2在相应位置从头建立DNA甲基化, 从而实现基因沉默(Erdmann and Picard, 2020).在杨柳科植物中发现1个促进雌性、抑制雄性的基因ARR17 (Müller et al., 2020; Xue et al., 2020).然而, 在一些杨属物种, 如欧洲山杨、毛果杨、美洲黑杨和胡杨(P. euphratica)中, 该基因并不位于Y染色体的性别决定区域(Y-SDR) (Yang et al., 2021).相应地, 在Y-SDR区域存在ARR17基因的反向重复序列(ARR17-IR).ARR17-IR可以转录产生长度为24 nt的siRNAs, 进一步通过RdDM途径引起ARR17启动子区域发生甲基化, 从而抑制ARR17的表达(图2).在柳属中的腺柳和钻天柳中也发现Y- SDR存在ARR17-IR片段, 表明ARR17-IR通过RdDM途径调控ARR17基因表达的性别决定方式在杨柳科植物中具有一定的保守性(Wang et al., 2022). ... ACC oxidase and miRNA- 159a, and their involvement in fresh fruit bunch yield (FFB) via sex ratio determination in oil palm 1 2016 ... MiRNA是一类长度为20-24 nt的非编码RNA, 可通过碱基互补配对的方式与靶基因mRNA结合, 使靶基因mRNA降解或者抑制其翻译, 实现miRNA对靶基因的负调控(Axtell, 2013).前期研究发现, miR172和miR169可控制花发育ABC类基因的表达, 进而控制矮牵牛(Petunia hybrida)和金鱼草(Antirrhinum majus)的花器官表型(Cartolano et al., 2007).近期, 有关雌雄异株植物的研究也发现miRNA在雌、雄个体间差异表达.例如, 在番木瓜花中共检测到65个miRNA, 其中14个miRNA在雌、雄花间差异表达(李崇奇, 2014).在油棕(Elaeis guineensis)中, miRNA159a表现出雌性偏向表达式样, 可能参与雌蕊发育(Somyong et al., 2016).对中华猕猴桃雌、雄花的小RNA组学分析显示, 170个miRNA在性别间差异表达, 并且在猕猴桃的性染色体(即Chr. 25)上预测到3个miRNA, 其中novel-ach-miR362的靶基因Achn298021与猕猴桃的单性花发育有关(闫明科等, 2015).此外, 对杜仲(Eucommia ulmoides)花芽进行小RNA测序, 共鉴定到187个miRNA, 其中31个为性别间差异表达的miRNA.进一步的靶基因预测分析显示, novel-23-miRNA调控花发育ABC类基因DEF的表达, 可能参与杜仲的性别决定(朱利利, 2019). ... Genome sequencing of the staple food crop white Guinea yam enables the development of a molecular marker for sex determination 1 2017 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Genetic and physical maps around the sex-determining M-locus of the dioecious plant asparagus 1 2007 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Repeated translocation of a gene cassette drives sex-chromosome turnover in strawberries 1 2018 ... Sex determination genes of dioecious plants
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Digital transcriptome analysis of putative sex-determination genes in papaya (Carica papaya) 1 2012 ... Sex determination genes of dioecious plants
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Repeated turnovers keep sex chromosomes young in willows 3 2022 ... Sex determination genes of dioecious plants
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... Genomes of the banyan tree and pollinator wasp provide insights into fig-wasp coevolution 2 2020 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ... The population genetics of structural variants in grapevine domestication 2 2019 ... Sex chromosome type of dioecious plants with reference genome
植物性别决定基因位于性染色体上的性别决定区域, 是雌雄性别分化的分子基础.从2014年第1个植物性别决定基因OGI被克隆, 至今已鉴定出23种雌雄异株植物的性别决定基因, 揭示了植物性别决定的“双基因模型”和“单基因模型”(表2).其中“双基因模型”是解释雌雄异株植物性别决定的经典理论, 该理论认为Y染色体上的性别决定区域存在2个连锁遗传的促雄基因(M)和抑雌基因(SuF) (Charlesworth and Charlesworth, 1978).在雄性个体中, M和SuF共同作用, 产生雄花; 而雌性个体由于缺乏这2个基因, 因此产生雌花.Akagi等(2018, 2019)通过全基因组测序、转录组分析和基因功能研究, 发现了猕猴桃Y染色体上性别决定区域(Y-SDR)的2个基因, 将其命名为抑雌基因SyGl和促雄基因FrBy.SyGl抑制雌蕊发育且FrBy促进雄蕊发育, 二者共同决定猕猴桃的性别.Harkess等(2017, 2020)基于基因组和转录组数据, 并结合诱变筛选与基因功能分析, 成功鉴定出芦笋的抑雌基因SOFF和促雄基因AspTDF1.Massonnet等(2020)分析了葡萄属(Vitis) 20个物种的Y-SDR区域序列, 并结合基因表达分析, 确定VviINP1基因控制葡萄的雄性育性.Iocco-Corena等(2021)进一步开展CRISPR/Cas基因编辑分析, 发现VviPLATZ1决定葡萄的雌性育性.在野生葡萄(V. vinifera subsp. sylvestris)驯化过程中, Y-SDR区域发生过1次罕见的重组事件, 产生了两性花突变体, 从而培育出现代的栽培葡萄(V. vinifera subsp. vinifera) (Zhou et al., 2019; Zou et al., 2021). ...
本文的其它图/表
|

 首页
首页