|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
硝酸盐转运蛋白NRT2在植物中的功能及分子机制研究进展
植物学报
2023, 58 (5):
783-798.
DOI: 10.11983/CBB22134
氮素作为植物生长发育所需的大量元素, 对植物生长发育及作物产量具有重要作用。施入氮肥是植物及作物的主要氮素来源。面对当下过度施肥造成面源污染加剧的现状, 提高作物氮素利用效率, 实现“减肥增产”的绿色增产增效模式, 是促进我国农业可持续发展及保障国家粮食安全的重要措施。当土壤氮匮缺时, 硝酸盐转运蛋白NRT2家族成员对根系吸收及转运硝酸盐至关重要, 其中NRT2.1在植物缺氮时主要负责根部的硝酸根吸收。该文重点总结了模式植物拟南芥(Arabidopsis thaliana)及重要粮油作物中NRT2家族蛋白特别是NRT2.1的功能及调控机理研究进展, 旨在为后续挖掘NRT2在提高作物产量方面的潜力及分子调控机制研究提供重要依据。
表2
参与调控NRT2的转录因子
正文中引用本图/表的段落
迄今为止, 在拟南芥中共鉴定到7个NRT2家族成员, 其中AtNRT2.1、AtNRT2.2、AtNRT2.4和AtNRT2.5属于高亲和硝酸盐转运系统(Orsel et al., 2002; Wang et al., 2018) (表1)。根系是植物吸收营养物质的主要器官, AtNRT2家族中的AtNRT2.1、AtNRT2.2、AtNRT2.4和AtNRT2.5均在根中表达。研究发现, 在1 mmol?L-1 NO3-处理下, 相较于AtNRT2.1和AtNRT- 2.2受到NO3-强烈诱导, AtNRT2.4在根中只有少量诱导, AtNRT2.5的表达则被NO3-强烈抑制(Okamoto et al., 2003)。在严重氮饥饿条件下, AtNRT2.4被大量诱导表达, 一旦外源供给氮素, 其表达水平会迅速下降(Okamoto et al., 2003; Kiba et al., 2012)。基于NO3-吸收的生理及表型实验证明, AtNRT2.1和AtNRT2.2在iHATS系统中发挥关键作用(Orsel et al., 2004)。进一步的遗传学实验表明, AtNRT2.1在iHATS中占主导作用, AtNRT2.2则起辅助吸收作用。在Atnrt2.1/ Atnrt2.2双突变体中回补AtNRT2.1, 其NO3-吸收恢复至野生型水平, 而当AtNRT2.1缺失后, 回补AtNRT- 2.2只能部分补偿植株的生长缺陷(Li et al., 2007)。与AtNRT2.1和AtNRT2.2不同, AtNRT2.4和AtNRT2.5只有在极低浓度的NO3-或严重氮饥饿条件下行使功能。在爪蟾(Xenopus laevis)卵母细胞中的实验结果显示, AtNRT2.4属于高亲和硝酸根转运蛋白, Atnrt2.4突变体在氮匮乏条件下氮吸收减少且地上部的氮含量也明显下降, 在AtNRT2.1及AtNRT2.2双突变的基础上突变AtNRT2.4 (三重突变体)后, 因根部硝酸根内流的减少对低氮下地上部的生物量产生更显著的影响(Kiba et al., 2012)。AtNRT2.5的表达同样受到氮饥饿诱导, 在长期遭受氮胁迫处理时, AtNRT2.5在拟南芥地上部及地下部根系中均被诱导表达(Lezhneva et al., 2014)。AtNRT2.4和AtNRT2.5还在茎的维管组织中表达, 其编码蛋白介导韧皮部NO3-的装载(Kiba et al., 2012; Lezhneva et al., 2014), 这表明AtNRT2.4和AtNRT2.5虽与AtNRT2.1及AtNRT2.2一样也介导根部的硝酸根吸收, 但其也参与硝酸根自根部向地上各组织的转运, 而且AtNRT2.5可能更多是在遭受氮饥饿的成年材料中发挥作用。
迄今为止, 在拟南芥中共鉴定到7个NRT2家族成员, 其中AtNRT2.1、AtNRT2.2、AtNRT2.4和AtNRT2.5属于高亲和硝酸盐转运系统(Orsel et al., 2002; Wang et al., 2018) (表1)。根系是植物吸收营养物质的主要器官, AtNRT2家族中的AtNRT2.1、AtNRT2.2、AtNRT2.4和AtNRT2.5均在根中表达。研究发现, 在1 mmol?L-1 NO3-处理下, 相较于AtNRT2.1和AtNRT- 2.2受到NO3-强烈诱导, AtNRT2.4在根中只有少量诱导, AtNRT2.5的表达则被NO3-强烈抑制(Okamoto et al., 2003)。在严重氮饥饿条件下, AtNRT2.4被大量诱导表达, 一旦外源供给氮素, 其表达水平会迅速下降(Okamoto et al., 2003; Kiba et al., 2012)。基于NO3-吸收的生理及表型实验证明, AtNRT2.1和AtNRT2.2在iHATS系统中发挥关键作用(Orsel et al., 2004)。进一步的遗传学实验表明, AtNRT2.1在iHATS中占主导作用, AtNRT2.2则起辅助吸收作用。在Atnrt2.1/ Atnrt2.2双突变体中回补AtNRT2.1, 其NO3-吸收恢复至野生型水平, 而当AtNRT2.1缺失后, 回补AtNRT- 2.2只能部分补偿植株的生长缺陷(Li et al., 2007)。与AtNRT2.1和AtNRT2.2不同, AtNRT2.4和AtNRT2.5只有在极低浓度的NO3-或严重氮饥饿条件下行使功能。在爪蟾(Xenopus laevis)卵母细胞中的实验结果显示, AtNRT2.4属于高亲和硝酸根转运蛋白, Atnrt2.4突变体在氮匮乏条件下氮吸收减少且地上部的氮含量也明显下降, 在AtNRT2.1及AtNRT2.2双突变的基础上突变AtNRT2.4 (三重突变体)后, 因根部硝酸根内流的减少对低氮下地上部的生物量产生更显著的影响(Kiba et al., 2012)。AtNRT2.5的表达同样受到氮饥饿诱导, 在长期遭受氮胁迫处理时, AtNRT2.5在拟南芥地上部及地下部根系中均被诱导表达(Lezhneva et al., 2014)。AtNRT2.4和AtNRT2.5还在茎的维管组织中表达, 其编码蛋白介导韧皮部NO3-的装载(Kiba et al., 2012; Lezhneva et al., 2014), 这表明AtNRT2.4和AtNRT2.5虽与AtNRT2.1及AtNRT2.2一样也介导根部的硝酸根吸收, 但其也参与硝酸根自根部向地上各组织的转运, 而且AtNRT2.5可能更多是在遭受氮饥饿的成年材料中发挥作用。
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al., 2007)。目前, 关于AtNRT2.3的功能还未见报道, 仅停留在不同组织及低浓度硝酸盐诱导的表达模式分析上, AtNRT2.3在根部的表达基本不受硝酸盐影响, 而且AtNRT2.3在不同营养生长期的拟南芥材料中具有差异化组织表达特征(Okamoto et al., 2003), 这在一定程度上暗示AtNRT2.3具有特殊功能。虽然AtNRT2.6未被检测到能够转运NO3-, 但是AtNRT2.6可能也与AtNRT2.1一样需要在伴侣蛋白的帮助下才具备硝酸根转运活性(Dechorgnat et al., 2012)。有意思的是, AtNRT2.6的转录受到高浓度硝酸盐和病原菌梨火疫病菌(Erwinia amylovora)的显著诱导, 并且其表达与活性氧(reactive oxygen species, ROS)的积累存在正相关, 由此也导致AtNRT2.6缺失突变体Atnrt2.6中活性氧含量减少, 同时表现出对梨火疫病菌的攻击更加敏感, 暗示AtNRT2.6可能介导植物对病原菌侵染的响应过程, 但其通过转运硝酸根还是其它分子介导该过程尚不清楚(Dechorgnat et al., 2012)。
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al., 2007)。目前, 关于AtNRT2.3的功能还未见报道, 仅停留在不同组织及低浓度硝酸盐诱导的表达模式分析上, AtNRT2.3在根部的表达基本不受硝酸盐影响, 而且AtNRT2.3在不同营养生长期的拟南芥材料中具有差异化组织表达特征(Okamoto et al., 2003), 这在一定程度上暗示AtNRT2.3具有特殊功能。虽然AtNRT2.6未被检测到能够转运NO3-, 但是AtNRT2.6可能也与AtNRT2.1一样需要在伴侣蛋白的帮助下才具备硝酸根转运活性(Dechorgnat et al., 2012)。有意思的是, AtNRT2.6的转录受到高浓度硝酸盐和病原菌梨火疫病菌(Erwinia amylovora)的显著诱导, 并且其表达与活性氧(reactive oxygen species, ROS)的积累存在正相关, 由此也导致AtNRT2.6缺失突变体Atnrt2.6中活性氧含量减少, 同时表现出对梨火疫病菌的攻击更加敏感, 暗示AtNRT2.6可能介导植物对病原菌侵染的响应过程, 但其通过转运硝酸根还是其它分子介导该过程尚不清楚(Dechorgnat et al., 2012)。
HATS基因的上调表达和根系构型的改变是植物应对土壤中氮匮乏时的两种适应策略, 当外界氮浓度较低时, 会显著促进植物侧根数目的增加及侧根的伸长以扩大获取氮素的根表面积(Zhang and Forde, 2000; Remans et al., 2006)。在植物根系对氮匮缺的综合响应中, 硝酸盐转运系统与根系构型调控存在相互作用(Remans et al., 2006)。拟南芥根系构型在应答不同浓度氮匮缺时表现有所不同, 当将拟南芥野生型Ws (Wassilewskija)从高氮(10 mmol?L-1 NO3-)转移至中度氮匮乏(1或0.5 mmol?L-1 NO3-)培养基时, 可见其侧根数目明显增加, 而当转至严重氮匮乏(0.1或0.05 mmol?L-1 NO3-)培养基中时, 侧根的平均长度显著增加(Remans et al., 2006)。研究发现, AtNRT2.1在根系对低氮胁迫的综合形态和生理响应中发挥至关重要的作用, 当拟南芥Atnrt2.1-1突变体在中度低氮条件下(0.5 mmol?L-1 NO3-)生长时, 与野生型表现不同的是其侧根数目无明显增加, 即Atnrt2.1突变导致缺氮时侧根减少, 但其侧根平均长度却显著增加, 表明Atnrt2.1突变体在中度缺氮的条件下表现出野生型拟南芥在极度氮匮乏时的根系构型, 这在一定程度上可以通过突变体中氮吸收活力减弱来解释, 同时也说明AtNRT2.1突变抑制氮匮缺下侧根发生并不依赖于根系的硝酸根吸收活力(Remans et al., 2006)。上述结果表明, AtNRT2.1在拟南芥响应外界氮水平时的根系构型改变过程中可能具有双重作用, 其一是作为高亲和硝酸盐转运系统的重要转运蛋白间接调控NO3-依赖的RSA响应; 其二则是通过目前尚不明确的特定功能直接调控氮匮缺时侧根的发生(Remans et al., 2006)。
HATS基因的上调表达和根系构型的改变是植物应对土壤中氮匮乏时的两种适应策略, 当外界氮浓度较低时, 会显著促进植物侧根数目的增加及侧根的伸长以扩大获取氮素的根表面积(Zhang and Forde, 2000; Remans et al., 2006)。在植物根系对氮匮缺的综合响应中, 硝酸盐转运系统与根系构型调控存在相互作用(Remans et al., 2006)。拟南芥根系构型在应答不同浓度氮匮缺时表现有所不同, 当将拟南芥野生型Ws (Wassilewskija)从高氮(10 mmol?L-1 NO3-)转移至中度氮匮乏(1或0.5 mmol?L-1 NO3-)培养基时, 可见其侧根数目明显增加, 而当转至严重氮匮乏(0.1或0.05 mmol?L-1 NO3-)培养基中时, 侧根的平均长度显著增加(Remans et al., 2006)。研究发现, AtNRT2.1在根系对低氮胁迫的综合形态和生理响应中发挥至关重要的作用, 当拟南芥Atnrt2.1-1突变体在中度低氮条件下(0.5 mmol?L-1 NO3-)生长时, 与野生型表现不同的是其侧根数目无明显增加, 即Atnrt2.1突变导致缺氮时侧根减少, 但其侧根平均长度却显著增加, 表明Atnrt2.1突变体在中度缺氮的条件下表现出野生型拟南芥在极度氮匮乏时的根系构型, 这在一定程度上可以通过突变体中氮吸收活力减弱来解释, 同时也说明AtNRT2.1突变抑制氮匮缺下侧根发生并不依赖于根系的硝酸根吸收活力(Remans et al., 2006)。上述结果表明, AtNRT2.1在拟南芥响应外界氮水平时的根系构型改变过程中可能具有双重作用, 其一是作为高亲和硝酸盐转运系统的重要转运蛋白间接调控NO3-依赖的RSA响应; 其二则是通过目前尚不明确的特定功能直接调控氮匮缺时侧根的发生(Remans et al., 2006)。
此外, 根系中硝酸盐的积累还与根系水分运输密切相关。当AtNRT2.1突变后, 根系的水分运输能力降低30%, 表明AtNRT2.1对根系导水率和质膜水通道蛋白活性具有调控作用。相关数据显示, AtNRT2.1对根系导水率的调控不依赖硝酸根(Li et al., 2016), 表明AtNRT2.1可能在提升植株抗旱性以及其它与根系导水率相关的生物学过程中发挥重要作用。
此外, 根系中硝酸盐的积累还与根系水分运输密切相关。当AtNRT2.1突变后, 根系的水分运输能力降低30%, 表明AtNRT2.1对根系导水率和质膜水通道蛋白活性具有调控作用。相关数据显示, AtNRT2.1对根系导水率的调控不依赖硝酸根(Li et al., 2016), 表明AtNRT2.1可能在提升植株抗旱性以及其它与根系导水率相关的生物学过程中发挥重要作用。
NRT2家族成员在水稻中的功能研究也取得了一系列进展。研究表明, 过表达OsNRT2.1使得水稻种子根、不定根和侧根总长度增加, 且在OsNRT2.1过表达植株中生长素转运蛋白基因OsPIN1a/b/c和OsPIN2的表达量也显著升高, 暗示OsNRT2.1可能通过调控生长素向根系的转运参与硝酸盐依赖的根系伸长过程(Naz et al., 2019), 但过量表达OsNRT- 2.1并未增加NO3-的吸收速率(Katayama et al., 2009)。在水稻中利用Ubiquitin启动子过表达OsNRT2.1, 尽管总分蘖数、生物量及总含氮量均显著提高, 但由于缺少伴侣蛋白OsNAR2.1的同步提高, 最终导致其由茎秆及叶向穗中的氮转运效率明显下降, 从而导致单株产量显著降低(陈景光等, 2016; Chen et al., 2016a), 这与Katayama等(2009)的报道相似, 也再次证明OsNRT2.1与其伴侣蛋白OsNAR2.1的结合对其发挥吸收硝酸盐功能的重要性(Feng et al., 2011)。有意思的是, 当以OsNAR2.1的启动子驱动OsNRT2.1 (pOsNAR2.1:OsNRT2.1)在水稻中过量表达时, 能显著促进转基因株系的生长, 提高氮素吸收效率及籽粒产量, 其农学氮素利用效率(agricultural nitrogen- use efficiency, ANUE)是野生型的128% (Chen et al., 2016a; 陈景光, 2017)。
NRT2家族成员在水稻中的功能研究也取得了一系列进展。研究表明, 过表达OsNRT2.1使得水稻种子根、不定根和侧根总长度增加, 且在OsNRT2.1过表达植株中生长素转运蛋白基因OsPIN1a/b/c和OsPIN2的表达量也显著升高, 暗示OsNRT2.1可能通过调控生长素向根系的转运参与硝酸盐依赖的根系伸长过程(Naz et al., 2019), 但过量表达OsNRT- 2.1并未增加NO3-的吸收速率(Katayama et al., 2009)。在水稻中利用Ubiquitin启动子过表达OsNRT2.1, 尽管总分蘖数、生物量及总含氮量均显著提高, 但由于缺少伴侣蛋白OsNAR2.1的同步提高, 最终导致其由茎秆及叶向穗中的氮转运效率明显下降, 从而导致单株产量显著降低(陈景光等, 2016; Chen et al., 2016a), 这与Katayama等(2009)的报道相似, 也再次证明OsNRT2.1与其伴侣蛋白OsNAR2.1的结合对其发挥吸收硝酸盐功能的重要性(Feng et al., 2011)。有意思的是, 当以OsNAR2.1的启动子驱动OsNRT2.1 (pOsNAR2.1:OsNRT2.1)在水稻中过量表达时, 能显著促进转基因株系的生长, 提高氮素吸收效率及籽粒产量, 其农学氮素利用效率(agricultural nitrogen- use efficiency, ANUE)是野生型的128% (Chen et al., 2016a; 陈景光, 2017)。
与其它OsNRT2成员不同, 水稻OsNRT2.4在爪蟾卵母细胞体系中被证明具有双亲和硝酸盐转运活性, 其蛋白高亲和力KM值为0.15 mmol?L-1, 低亲和力KM值为4 mmol?L-1 (Wei et al., 2018)。据报道, OsNRT2.4参与调控侧根生长, 在水稻根中, OsNRT2.4主要在侧根原基基部表达, 该基因敲除后显著降低了侧根数目和侧根长度。与OsNRT2.1类似, OsNRT2.4对根系生长的调控被认为是通过其介导的硝酸盐转运功能来发挥作用(Wei et al., 2018)。在外界极低浓度NO3-条件下, OsNRT2.3a参与NO3-从根向地上部的长距离运输(Tang et al., 2012)。在水稻中同时过表达OsNRT2.3a及其伴侣蛋白OsNAR2.1可显著提高水稻产量及氮素利用效率(Chen et al., 2020)。另一成员OsNRT2.3b在感知韧皮部细胞胞质pH值以平衡NO3-和NH4+摄取中发挥关键作用(Fan et al., 2016)。过表达OsNRT2.3b对水稻生长、氮素利用效率及最终的籽粒产量均有所促进(陆海燕等, 2015; Fan et al., 2016), 说明水稻NRT2家族蛋白的硝酸根转运能力特殊且存在功能分化。
与其它OsNRT2成员不同, 水稻OsNRT2.4在爪蟾卵母细胞体系中被证明具有双亲和硝酸盐转运活性, 其蛋白高亲和力KM值为0.15 mmol?L-1, 低亲和力KM值为4 mmol?L-1 (Wei et al., 2018)。据报道, OsNRT2.4参与调控侧根生长, 在水稻根中, OsNRT2.4主要在侧根原基基部表达, 该基因敲除后显著降低了侧根数目和侧根长度。与OsNRT2.1类似, OsNRT2.4对根系生长的调控被认为是通过其介导的硝酸盐转运功能来发挥作用(Wei et al., 2018)。在外界极低浓度NO3-条件下, OsNRT2.3a参与NO3-从根向地上部的长距离运输(Tang et al., 2012)。在水稻中同时过表达OsNRT2.3a及其伴侣蛋白OsNAR2.1可显著提高水稻产量及氮素利用效率(Chen et al., 2020)。另一成员OsNRT2.3b在感知韧皮部细胞胞质pH值以平衡NO3-和NH4+摄取中发挥关键作用(Fan et al., 2016)。过表达OsNRT2.3b对水稻生长、氮素利用效率及最终的籽粒产量均有所促进(陆海燕等, 2015; Fan et al., 2016), 说明水稻NRT2家族蛋白的硝酸根转运能力特殊且存在功能分化。
除了在NO3-吸收和转运以及调控根系生长发育中发挥作用, OsNRT2s的一些成员还通过协同其它营养元素的吸收增加水稻产量。例如, 在水稻中过表达OsNRT2.1能够显著促进水稻对锰的吸收, 在干湿交替栽培模式下OsNRT2.1的超量表达能够显著提高水稻产量(Luo et al., 2018); 过表达OsNRT2.3b则显著促进水稻对磷的吸收和积累, 并促进籽粒产量和秸秆生物量增加(Feng et al., 2017), 说明OsNRT2s通过协同其它大量或者微量元素的吸收来促进水稻生长, 进而提高产量。
除了在NO3-吸收和转运以及调控根系生长发育中发挥作用, OsNRT2s的一些成员还通过协同其它营养元素的吸收增加水稻产量。例如, 在水稻中过表达OsNRT2.1能够显著促进水稻对锰的吸收, 在干湿交替栽培模式下OsNRT2.1的超量表达能够显著提高水稻产量(Luo et al., 2018); 过表达OsNRT2.3b则显著促进水稻对磷的吸收和积累, 并促进籽粒产量和秸秆生物量增加(Feng et al., 2017), 说明OsNRT2s通过协同其它大量或者微量元素的吸收来促进水稻生长, 进而提高产量。
在禾本科作物玉米中, 目前已鉴定到ZmNRT2.1、Zm- NRT2.2、ZmNRT2.3和ZmNRT2.5共4个NRT2家族的硝酸根转运蛋白(Plett et al., 2010) (表1)。该家族中只有ZmNRT2.1和ZmNRT2.2两个基因亲缘关系较近, 氨基酸序列同源性达98%。ZmNRT2.1和ZmNRT2.2的转录水平在根部均受NO3-诱导, 原位杂交结果表明ZmNRT2.1特异性在皮层表达, 而ZmNRT2.2除分布于皮层, 还在中柱和侧根原基中表达(Trevisan et al., 2008), 并且玉米根系对NO3-吸收能力的增强与ZmNRT2.1和ZmNRT2.2转录水平的快速上调相对应(Sabermanesh et al., 2017)。ZmNRT2.5的表达仅在氮饥饿和低氮条件下被检测到, 并且受到氮饥饿(无氮)的强烈诱导, 在植株重新补充氮(2.5 mmol?L-1 NH4NO3)后, 根和茎中ZmNRT2.5的表达量均下调(Dechorgnat et al., 2019); 当玉米进入生殖生长期, ZmNRT2.5在根中的表达趋近于零, 反而在苞叶中表达量较高, 推测ZmNRT2.5可能在灌浆期籽粒氮分配中起重要作用(Fujita et al., 1995)。迄今为止, 关于ZmNRT2家族成员的相关研究基本都在转录水平, 还未见生理及遗传学实验验证ZmNRT2的NO3-转运活性以及亲和力。
在禾本科作物玉米中, 目前已鉴定到ZmNRT2.1、Zm- NRT2.2、ZmNRT2.3和ZmNRT2.5共4个NRT2家族的硝酸根转运蛋白(Plett et al., 2010) (表1)。该家族中只有ZmNRT2.1和ZmNRT2.2两个基因亲缘关系较近, 氨基酸序列同源性达98%。ZmNRT2.1和ZmNRT2.2的转录水平在根部均受NO3-诱导, 原位杂交结果表明ZmNRT2.1特异性在皮层表达, 而ZmNRT2.2除分布于皮层, 还在中柱和侧根原基中表达(Trevisan et al., 2008), 并且玉米根系对NO3-吸收能力的增强与ZmNRT2.1和ZmNRT2.2转录水平的快速上调相对应(Sabermanesh et al., 2017)。ZmNRT2.5的表达仅在氮饥饿和低氮条件下被检测到, 并且受到氮饥饿(无氮)的强烈诱导, 在植株重新补充氮(2.5 mmol?L-1 NH4NO3)后, 根和茎中ZmNRT2.5的表达量均下调(Dechorgnat et al., 2019); 当玉米进入生殖生长期, ZmNRT2.5在根中的表达趋近于零, 反而在苞叶中表达量较高, 推测ZmNRT2.5可能在灌浆期籽粒氮分配中起重要作用(Fujita et al., 1995)。迄今为止, 关于ZmNRT2家族成员的相关研究基本都在转录水平, 还未见生理及遗传学实验验证ZmNRT2的NO3-转运活性以及亲和力。
豆科植物的根系结瘤过程非常耗能, 一旦环境中氮素营养充足, 豆科植物会主动选择暂时终止根系结瘤。截至目前, 已报道的研究结果大多关注结瘤的“氮阻遏”机制, 而关于豆科植物如何感应外界氮水平进而精密地调控结瘤固氮的开启或关闭仍然是一个未解之谜。作为在氮匮乏条件下发挥主要功能的高亲和硝酸根转运系统也备受研究者关注。在既是豆科模式植物又是重要牧草的百脉根(Lotus japonicus)中的研究表明, NRT2家族成员参与高氮下的结瘤抑制过程。百脉根LjNRT2.1的功能缺失导致突变体的硝酸根吸收能力减弱, Ljnrt2.1突变体在高氮下仍然保持结瘤, 说明在高氮下LjNRT2.1可能通过介导硝酸盐吸收和转运来抑制结瘤(Misawa et al., 2022)。Valkov等(2020)则发现百脉根中的另一个NRT2成员LjNRT2.4在低氧条件下调控硝酸盐维持根瘤活性方面发挥作用, Ljnrt2.4突变体的根瘤中硝酸盐含量减少导致根瘤固氮酶活性明显下降, 同时在水培厌氧条件下植物也表现出重度氮匮缺表型, 地上部生物量较野生型显著降低。进一步的研究结果显示, LjNRT2.4缺失突变对根瘤中NO生物合成与积累也产生重要影响, 表明LjNRT2.4对百脉根的根瘤固氮具有促进作用(Valkov et al., 2020)。
豆科植物的根系结瘤过程非常耗能, 一旦环境中氮素营养充足, 豆科植物会主动选择暂时终止根系结瘤。截至目前, 已报道的研究结果大多关注结瘤的“氮阻遏”机制, 而关于豆科植物如何感应外界氮水平进而精密地调控结瘤固氮的开启或关闭仍然是一个未解之谜。作为在氮匮乏条件下发挥主要功能的高亲和硝酸根转运系统也备受研究者关注。在既是豆科模式植物又是重要牧草的百脉根(Lotus japonicus)中的研究表明, NRT2家族成员参与高氮下的结瘤抑制过程。百脉根LjNRT2.1的功能缺失导致突变体的硝酸根吸收能力减弱, Ljnrt2.1突变体在高氮下仍然保持结瘤, 说明在高氮下LjNRT2.1可能通过介导硝酸盐吸收和转运来抑制结瘤(Misawa et al., 2022)。Valkov等(2020)则发现百脉根中的另一个NRT2成员LjNRT2.4在低氧条件下调控硝酸盐维持根瘤活性方面发挥作用, Ljnrt2.4突变体的根瘤中硝酸盐含量减少导致根瘤固氮酶活性明显下降, 同时在水培厌氧条件下植物也表现出重度氮匮缺表型, 地上部生物量较野生型显著降低。进一步的研究结果显示, LjNRT2.4缺失突变对根瘤中NO生物合成与积累也产生重要影响, 表明LjNRT2.4对百脉根的根瘤固氮具有促进作用(Valkov et al., 2020)。
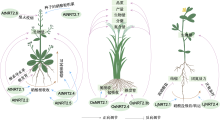
蓝色线条表示已经明确具有氮吸收及转运功能的NRT2家族成员; 棕色线条表示硝酸根吸收及转运; 蓝色虚线代表功能尚不明确; 紫色线条指示目前在拟南芥、水稻及百脉根中NRT2蛋白参与调控的其它生物学过程。
蓝色线条表示已经明确具有氮吸收及转运功能的NRT2家族成员; 棕色线条表示硝酸根吸收及转运; 蓝色虚线代表功能尚不明确; 紫色线条指示目前在拟南芥、水稻及百脉根中NRT2蛋白参与调控的其它生物学过程。
施加硝酸盐会强烈影响植物基因的表达, 根据植物对硝酸盐的应答, 可分为初级硝酸盐反应(primary nitrate response, PNR)和系统信号转导。PNR中的硝酸盐感知系统为组成型表达, 它不需要蛋白质的从头合成, 其下游基因或蛋白能够在几分钟内迅速响应硝酸盐信号(Ho et al., 2009; Medici and Krouk, 2014)。硝酸盐的系统信号转导则是植物通过感应体外或体内硝酸盐的状态及对氮的需求, 对地上-地下、细胞-细胞之间的氮信号进行交流和整合, 以调节硝酸盐吸收、同化、分配和植物发育(表2)。作为在根系吸收硝酸盐过程中起主要作用的AtNRT2.1, 其转录水平的调控备受关注。
施加硝酸盐会强烈影响植物基因的表达, 根据植物对硝酸盐的应答, 可分为初级硝酸盐反应(primary nitrate response, PNR)和系统信号转导。PNR中的硝酸盐感知系统为组成型表达, 它不需要蛋白质的从头合成, 其下游基因或蛋白能够在几分钟内迅速响应硝酸盐信号(Ho et al., 2009; Medici and Krouk, 2014)。硝酸盐的系统信号转导则是植物通过感应体外或体内硝酸盐的状态及对氮的需求, 对地上-地下、细胞-细胞之间的氮信号进行交流和整合, 以调节硝酸盐吸收、同化、分配和植物发育(表2)。作为在根系吸收硝酸盐过程中起主要作用的AtNRT2.1, 其转录水平的调控备受关注。
在植物根系对硝酸盐的级联响应中, NLP6和NLP7是主要的转录调控因子(Konishi and Yanagisawa, 2013)。研究发现, 拟南芥NLP6活性被抑制导致NRT1.1和NRT2.1等基因下调表达(Konishi and Yanagisawa, 2013); 同样, 过表达NLP7也会导致一系列涉及硝酸盐转运(NRT1.1和NRT2.1)、氮同化(GS1、NIA1、NIA2和NIR1)以及氮信号通路(LBD37、LBD38、LBD39、ANR1和AFB3)相关基因上调表达(Yu et al., 2016)。Marchive等(2013)研究发现, NO3-诱导NLP7从胞质向细胞核迁移并在核中积累, 利用在nlp7-1突变体背景下表达pNLP7:NLP7-GFP的材料, 施加NO3-诱导10分钟后进行染色质免疫共沉淀, 通过染色质免疫共沉淀-芯片(ChIP-chip)技术高通量筛选到NLP7结合的靶基因851个, 其中包括AtNRT2.1和AtNRT2.2; 接着又通过定量ChIP-PCR, 以及在原生质体中利用pNRT2.1:LUC和p35S:NLP7共转进行瞬时反式激活实验, 充分验证了NLP7可以直接与NRT2.1的启动子区结合, 进而调控早期硝酸盐应答。相关研究结果表明, NLPs可以靶向NRT2.1调控其表达, 在NO3-的早期响应过程中发挥重要作用。
在植物根系对硝酸盐的级联响应中, NLP6和NLP7是主要的转录调控因子(Konishi and Yanagisawa, 2013)。研究发现, 拟南芥NLP6活性被抑制导致NRT1.1和NRT2.1等基因下调表达(Konishi and Yanagisawa, 2013); 同样, 过表达NLP7也会导致一系列涉及硝酸盐转运(NRT1.1和NRT2.1)、氮同化(GS1、NIA1、NIA2和NIR1)以及氮信号通路(LBD37、LBD38、LBD39、ANR1和AFB3)相关基因上调表达(Yu et al., 2016)。Marchive等(2013)研究发现, NO3-诱导NLP7从胞质向细胞核迁移并在核中积累, 利用在nlp7-1突变体背景下表达pNLP7:NLP7-GFP的材料, 施加NO3-诱导10分钟后进行染色质免疫共沉淀, 通过染色质免疫共沉淀-芯片(ChIP-chip)技术高通量筛选到NLP7结合的靶基因851个, 其中包括AtNRT2.1和AtNRT2.2; 接着又通过定量ChIP-PCR, 以及在原生质体中利用pNRT2.1:LUC和p35S:NLP7共转进行瞬时反式激活实验, 充分验证了NLP7可以直接与NRT2.1的启动子区结合, 进而调控早期硝酸盐应答。相关研究结果表明, NLPs可以靶向NRT2.1调控其表达, 在NO3-的早期响应过程中发挥重要作用。
在豆科植物的根系结瘤过程中, NRT2.1也被发现受到NLP家族转录因子的调控。研究人员发现在Ljnlp1突变体中, NO3-对LjNRT2.1的诱导作用被显著抑制, 但是在Ljnlp4突变体中不受影响。反式激活实验发现, LjNLP1直接结合LjNRT2.1的启动子区以响应硝酸盐水平, 说明LjNRT2.1和LjNLP1共同参与硝酸盐诱导的结瘤调控和NO3-转运过程。进一步的研究表明, LjNLP4受NO3-诱导的核定位特征依赖于LjNLP1- LjNRT2.1模块, 即LjNLP4作用于LjNLP1-LjNRT2.1的下游(Misawa et al., 2022)。因此, LjNRT2.1介导的结瘤调控机制可以通过以下模型(LjNLP1-LjNRT2.1- LjNLP4信号通路的模型)得以阐明: 硝酸盐(10 mmol?L-1 KNO3)激活LjNLP1, 从而诱导LjNRT2.1的表达, 促进根系对硝酸盐的吸收及转运, 而NO3-的内流增强触发了LjNLP4的核定位, LjNLP4则诱导结瘤负调控因子LjCLE-RS2的表达。此外, LjNLP4还通过干扰百脉根中LjNIN (NODULE INCEPTION)的功能来抑制结瘤, 正调控基因的表达(Misawa et al., 2022)。上述结果表明, NLP家族转录因子对NRT家族成员特别是NRT2家族成员的靶向调控具有保守性。
在豆科植物的根系结瘤过程中, NRT2.1也被发现受到NLP家族转录因子的调控。研究人员发现在Ljnlp1突变体中, NO3-对LjNRT2.1的诱导作用被显著抑制, 但是在Ljnlp4突变体中不受影响。反式激活实验发现, LjNLP1直接结合LjNRT2.1的启动子区以响应硝酸盐水平, 说明LjNRT2.1和LjNLP1共同参与硝酸盐诱导的结瘤调控和NO3-转运过程。进一步的研究表明, LjNLP4受NO3-诱导的核定位特征依赖于LjNLP1- LjNRT2.1模块, 即LjNLP4作用于LjNLP1-LjNRT2.1的下游(Misawa et al., 2022)。因此, LjNRT2.1介导的结瘤调控机制可以通过以下模型(LjNLP1-LjNRT2.1- LjNLP4信号通路的模型)得以阐明: 硝酸盐(10 mmol?L-1 KNO3)激活LjNLP1, 从而诱导LjNRT2.1的表达, 促进根系对硝酸盐的吸收及转运, 而NO3-的内流增强触发了LjNLP4的核定位, LjNLP4则诱导结瘤负调控因子LjCLE-RS2的表达。此外, LjNLP4还通过干扰百脉根中LjNIN (NODULE INCEPTION)的功能来抑制结瘤, 正调控基因的表达(Misawa et al., 2022)。上述结果表明, NLP家族转录因子对NRT家族成员特别是NRT2家族成员的靶向调控具有保守性。
近年来, 研究人员陆续鉴定到多个转录因子参与调控NRT2.1。其中, 负调控因子NIGT1 (NITRATE- INDUCIBLE GARP-type TRANSCRIPTIONAL REPRESSOR 1)是一类由NO3-诱导的转录抑制子。研究表明NIGT1.1可以在低氮胁迫下抑制NLP7驱动的AtNRT2.1表达的激活, 且通过凝胶迁移(EMSA)实验证明NIGT1.1可以直接结合AtNRT2.1的启动子区; 此外, 共转染实验结果显示, 与NIGT1.1属于同一支系的同源基因(包括NIGT1.2、NIGT1.3和NIGT1.4)均受NO3-诱导, 并且在抑制硝酸盐依赖的AtNRT2.1表达激活中存在功能冗余(Maeda et al., 2018)。值得注意的是, 磷饥饿反应的主调控因子PHR1 (PHOSPHATE STARVATION RESPONSE 1)可以直接激活NIGT1s基因的表达, 导致硝酸盐的摄取减少(Maeda et al., 2018), 表明磷饥饿可能通过PHR1- NIGT1-NRT2.1的转录级联影响硝酸根的吸收。相反, 属于bZIP (BASIC REGION/LEUCINE ZIPPER)转录因子家族的TGA1 (TGACG MOTIF-BINDING FACTOR 1)和TGA4则对NRT2家族基因发挥正向调控作用(Alvarez et al., 2014)。在tag1/4双突变体中, NO3-诱导的AtNRT2.1和AtNRT2.2基因表达量显著降低; 同时, 在原生质体中过表达TGA1或TGA4, AtNRT2.1和AtNRT2.2基因的表达量明显高于对照组, 表明TGA1和TGA4正向调控AtNRT2.1和AtNRT2.2的表达(Alvarez et al., 2014)。进一步通过ChIP和酵母单杂交实验证明, TGA1可以直接结合AtNRT2.1和AtNRT2.2的启动子区(Alvarez et al., 2014)。反向遗传学研究证明, 侧生器官边界结构域蛋白LBD37/ 38/39也是调控硝酸盐应答反应中重要的转录因子, 并受NO3-强烈诱导。LBD37/38/39的过表达可以抑制AtNRT2.1、AtNRT2.2以及氮同化相关基因的表达, 表明LBD的这3个成员在硝酸盐信号转导中起负调控作用(Rubin et al., 2009), 但是目前尚未证实LBD是否能够直接靶向调控NRT2家族成员的表达。
近年来, 研究人员陆续鉴定到多个转录因子参与调控NRT2.1。其中, 负调控因子NIGT1 (NITRATE- INDUCIBLE GARP-type TRANSCRIPTIONAL REPRESSOR 1)是一类由NO3-诱导的转录抑制子。研究表明NIGT1.1可以在低氮胁迫下抑制NLP7驱动的AtNRT2.1表达的激活, 且通过凝胶迁移(EMSA)实验证明NIGT1.1可以直接结合AtNRT2.1的启动子区; 此外, 共转染实验结果显示, 与NIGT1.1属于同一支系的同源基因(包括NIGT1.2、NIGT1.3和NIGT1.4)均受NO3-诱导, 并且在抑制硝酸盐依赖的AtNRT2.1表达激活中存在功能冗余(Maeda et al., 2018)。值得注意的是, 磷饥饿反应的主调控因子PHR1 (PHOSPHATE STARVATION RESPONSE 1)可以直接激活NIGT1s基因的表达, 导致硝酸盐的摄取减少(Maeda et al., 2018), 表明磷饥饿可能通过PHR1- NIGT1-NRT2.1的转录级联影响硝酸根的吸收。相反, 属于bZIP (BASIC REGION/LEUCINE ZIPPER)转录因子家族的TGA1 (TGACG MOTIF-BINDING FACTOR 1)和TGA4则对NRT2家族基因发挥正向调控作用(Alvarez et al., 2014)。在tag1/4双突变体中, NO3-诱导的AtNRT2.1和AtNRT2.2基因表达量显著降低; 同时, 在原生质体中过表达TGA1或TGA4, AtNRT2.1和AtNRT2.2基因的表达量明显高于对照组, 表明TGA1和TGA4正向调控AtNRT2.1和AtNRT2.2的表达(Alvarez et al., 2014)。进一步通过ChIP和酵母单杂交实验证明, TGA1可以直接结合AtNRT2.1和AtNRT2.2的启动子区(Alvarez et al., 2014)。反向遗传学研究证明, 侧生器官边界结构域蛋白LBD37/ 38/39也是调控硝酸盐应答反应中重要的转录因子, 并受NO3-强烈诱导。LBD37/38/39的过表达可以抑制AtNRT2.1、AtNRT2.2以及氮同化相关基因的表达, 表明LBD的这3个成员在硝酸盐信号转导中起负调控作用(Rubin et al., 2009), 但是目前尚未证实LBD是否能够直接靶向调控NRT2家族成员的表达。
NRT2.1除了在初级硝酸盐反应中受上述转录因子的调控, 在硝酸盐介导的系统性信号通路中, NRT2.1可能作为潜在的调控枢纽。研究表明, 拟南芥转录因子TCP20 (TEOSINATE BRANCHED 1/CYC-LOIDEA/ PROLIFERATING CELL FACTOR 20)调控低硝酸盐条件下拟南芥根部觅食的系统信号转导, 体内和体外的研究结果显示TCP20可以与AtNRT2.1、AtNRT1.1以及AtNIA1 (NITRATE REDUCTASE NADH 1, NIA1)的启动子区直接结合(Guan et al., 2014)。然而, 在5 mmol?L-1 KNO3培养基上生长10天后分别检测野生型和tcp20突变体中AtNRT2.1、AtNRT1.1和AtNIA1的表达, 发现突变体中的AtNRT1.1和AtNIA1表达水平显著下降, 但是AtNRT2.1的表达量与野生型(WT)无明显差异; 通过分根实验也证实无论低氮(0 mmol?L-1 KNO3)还是高氮(5 mmol?L-1 KNO3), AtNRT2.1的表达量不受TCP20突变的影响(Guan et al., 2014)。因此, TCP20与AtNRT2.1的遗传学关系和转录调控机制还需深入研究。
NRT2.1除了在初级硝酸盐反应中受上述转录因子的调控, 在硝酸盐介导的系统性信号通路中, NRT2.1可能作为潜在的调控枢纽。研究表明, 拟南芥转录因子TCP20 (TEOSINATE BRANCHED 1/CYC-LOIDEA/ PROLIFERATING CELL FACTOR 20)调控低硝酸盐条件下拟南芥根部觅食的系统信号转导, 体内和体外的研究结果显示TCP20可以与AtNRT2.1、AtNRT1.1以及AtNIA1 (NITRATE REDUCTASE NADH 1, NIA1)的启动子区直接结合(Guan et al., 2014)。然而, 在5 mmol?L-1 KNO3培养基上生长10天后分别检测野生型和tcp20突变体中AtNRT2.1、AtNRT1.1和AtNIA1的表达, 发现突变体中的AtNRT1.1和AtNIA1表达水平显著下降, 但是AtNRT2.1的表达量与野生型(WT)无明显差异; 通过分根实验也证实无论低氮(0 mmol?L-1 KNO3)还是高氮(5 mmol?L-1 KNO3), AtNRT2.1的表达量不受TCP20突变的影响(Guan et al., 2014)。因此, TCP20与AtNRT2.1的遗传学关系和转录调控机制还需深入研究。
众所周知, 根系对硝酸盐的吸收受植物体内氮饱和的系统性反馈抑制, 从而调整根系对氮的需求。目前, 普遍认为这一机制涉及由韧皮部从茎向根传递的长距离信号, 如氮同化产物谷氨酰胺对根系NO3-吸收系统的反馈抑制(Cooper and Clarkson, 1989), 而NRT2.1被认为是这一反馈机制的主要调控靶点(Widiez et al., 2011)。如图2所示, Widiez等(2011)报道了1个编码拟南芥RNA聚合酶II复合体组分蛋白(AtIWS1) INTERACT WITH SPT6的基因HNI9 (HIGH NITROGEN INSENSITIVE 9), 在高氮处理下, HNI9/AtIWS1在根中抑制AtNRT2.1的转录, 初步研究显示, 高氮抑制AtNRT2.1的转录与AtNRT2.1位点组蛋白H3第27位赖氨酸的三甲基化修饰(H3K27me3)有关, 而HNI9/AtIWS1特异性参与H3K27me3的沉积。相关研究结果为揭示NRT2.1受高氮抑制的分子机制提供了新思路。近年来, 研究人员发现拟南芥光响应转录因子HY5 (ELONGATED HYPOCOTYL 5)可以从茎移动到根部, 并激活根部的HY5, 从而激活AtNRT2.1的表达, 进而促进根中硝酸盐的吸收(Chen et al., 2016b), 表明NRT2.1也可以接受来自地上部的信号因子。
众所周知, 根系对硝酸盐的吸收受植物体内氮饱和的系统性反馈抑制, 从而调整根系对氮的需求。目前, 普遍认为这一机制涉及由韧皮部从茎向根传递的长距离信号, 如氮同化产物谷氨酰胺对根系NO3-吸收系统的反馈抑制(Cooper and Clarkson, 1989), 而NRT2.1被认为是这一反馈机制的主要调控靶点(Widiez et al., 2011)。如图2所示, Widiez等(2011)报道了1个编码拟南芥RNA聚合酶II复合体组分蛋白(AtIWS1) INTERACT WITH SPT6的基因HNI9 (HIGH NITROGEN INSENSITIVE 9), 在高氮处理下, HNI9/AtIWS1在根中抑制AtNRT2.1的转录, 初步研究显示, 高氮抑制AtNRT2.1的转录与AtNRT2.1位点组蛋白H3第27位赖氨酸的三甲基化修饰(H3K27me3)有关, 而HNI9/AtIWS1特异性参与H3K27me3的沉积。相关研究结果为揭示NRT2.1受高氮抑制的分子机制提供了新思路。近年来, 研究人员发现拟南芥光响应转录因子HY5 (ELONGATED HYPOCOTYL 5)可以从茎移动到根部, 并激活根部的HY5, 从而激活AtNRT2.1的表达, 进而促进根中硝酸盐的吸收(Chen et al., 2016b), 表明NRT2.1也可以接受来自地上部的信号因子。
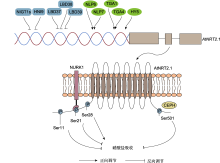
双螺旋表示AtNRT2.1的启动子区域, 受到多个转录因子的直接调控。其中蓝色矩形代表转录抑制子, 绿色椭圆形代表转录激活子。定位在质膜上的AtNRT2.1蛋白通过与NURK1激酶或CEPH磷酸酶的互作发生磷酸化或去磷酸化修饰。目前明确调控硝酸根吸收活性的磷酸化位点包括N端Ser11、Ser21、Ser28及C端Ser501。
双螺旋表示AtNRT2.1的启动子区域, 受到多个转录因子的直接调控。其中蓝色矩形代表转录抑制子, 绿色椭圆形代表转录激活子。定位在质膜上的AtNRT2.1蛋白通过与NURK1激酶或CEPH磷酸酶的互作发生磷酸化或去磷酸化修饰。目前明确调控硝酸根吸收活性的磷酸化位点包括N端Ser11、Ser21、Ser28及C端Ser501。
上述研究表明, AtNRT2.1转录水平的调控与硝酸盐的吸收及转运存在很强的相关性。然而, 当过表达AtNRT2.1时, 尽管AtNRT2.1转录本丰度积累, 但是植株HATS的活性在高氮下仍然受到抑制(Laugier et al., 2012), 暗示NRT2.1蛋白水平变化与其mRNA水平并不平行(Wirth et al., 2007)。大麦中的研究也发现, 在NO3-诱导过程中HvNRT2.1蛋白丰度的变化与HvNRT2.1转录本的积累或消减无明显关系(Ishikawa et al., 2009), 表明NRT2.1受到翻译后水平的调控。
上述研究表明, AtNRT2.1转录水平的调控与硝酸盐的吸收及转运存在很强的相关性。然而, 当过表达AtNRT2.1时, 尽管AtNRT2.1转录本丰度积累, 但是植株HATS的活性在高氮下仍然受到抑制(Laugier et al., 2012), 暗示NRT2.1蛋白水平变化与其mRNA水平并不平行(Wirth et al., 2007)。大麦中的研究也发现, 在NO3-诱导过程中HvNRT2.1蛋白丰度的变化与HvNRT2.1转录本的积累或消减无明显关系(Ishikawa et al., 2009), 表明NRT2.1受到翻译后水平的调控。
其次, 该领域的一些研究证明, 拟南芥AtNRT2.1蛋白的磷酸化和去磷酸化是调控其HATS活性的重要机制, 同时AtNRT2.1也是目前唯一被报道具有磷酸化修饰的NRT2家族成员(图2)。蛋白质磷酸化组学鉴定到拟南芥NRT2.1蛋白的N端和C端域(不包含跨膜域)存在磷酸化位点, 如Ser11、Ser21、Ser28、Ser501和Thr521, 不同位点的磷酸化水平随外界氮环境发生变化(Menz et al., 2016; Li et al., 2020; Jacquot et al., 2020)。低氮诱导拟南芥AtNRT2.1蛋白第28位丝氨酸残基(Ser28)磷酸化并使得蛋白的稳定性增强, 通过将模拟磷酸化失活的NRT2.1S28A以及组成型磷酸化状态的NRT2.1S28E回补拟南芥Atnrt2.1突变体, 发现Ser28位点失活突变导致其生长表型与Atnrt2.1突变体相似, 而NRT2.1S28E的地上部鲜重和NO3-吸收能力均恢复至野生型水平(Zou et al., 2020)。Li等(2020)同样证明AtNRT2.1蛋白Ser28的磷酸化促进NO3-内流, 与之相反, AtNRT2.1蛋白第11位丝氨酸残基(Ser11)磷酸化则起抑制作用, 表明Ser28是一个NO3-吸收的正调控位点, 而Ser11是一个负调控位点。此外, 另一个负调控位点AtNRT2.1蛋白第501位丝氨酸残基(Ser501)的磷酸化导致AtNRT2.1失活, 从而减少NO3-向根系的转运(Jacquot et al., 2020)。上述结果表明, 在感应外界不同环境的刺激下, NRT2.1蛋白不同位点的磷酸化具有正向或负向调节NO3-吸收的作用, 说明在翻译后水平NRT2.1受到了非常精密且复杂的调控。
其次, 该领域的一些研究证明, 拟南芥AtNRT2.1蛋白的磷酸化和去磷酸化是调控其HATS活性的重要机制, 同时AtNRT2.1也是目前唯一被报道具有磷酸化修饰的NRT2家族成员(图2)。蛋白质磷酸化组学鉴定到拟南芥NRT2.1蛋白的N端和C端域(不包含跨膜域)存在磷酸化位点, 如Ser11、Ser21、Ser28、Ser501和Thr521, 不同位点的磷酸化水平随外界氮环境发生变化(Menz et al., 2016; Li et al., 2020; Jacquot et al., 2020)。低氮诱导拟南芥AtNRT2.1蛋白第28位丝氨酸残基(Ser28)磷酸化并使得蛋白的稳定性增强, 通过将模拟磷酸化失活的NRT2.1S28A以及组成型磷酸化状态的NRT2.1S28E回补拟南芥Atnrt2.1突变体, 发现Ser28位点失活突变导致其生长表型与Atnrt2.1突变体相似, 而NRT2.1S28E的地上部鲜重和NO3-吸收能力均恢复至野生型水平(Zou et al., 2020)。Li等(2020)同样证明AtNRT2.1蛋白Ser28的磷酸化促进NO3-内流, 与之相反, AtNRT2.1蛋白第11位丝氨酸残基(Ser11)磷酸化则起抑制作用, 表明Ser28是一个NO3-吸收的正调控位点, 而Ser11是一个负调控位点。此外, 另一个负调控位点AtNRT2.1蛋白第501位丝氨酸残基(Ser501)的磷酸化导致AtNRT2.1失活, 从而减少NO3-向根系的转运(Jacquot et al., 2020)。上述结果表明, 在感应外界不同环境的刺激下, NRT2.1蛋白不同位点的磷酸化具有正向或负向调节NO3-吸收的作用, 说明在翻译后水平NRT2.1受到了非常精密且复杂的调控。
尽管有些磷酸化位点的功能已经明确, 但对于NRT2.1磷酸化修饰的发生及其作用机制还知之甚少。研究表明, NRT2.1与NAR2蛋白的互作强度可能与NRT2.1蛋白的磷酸化修饰有关。Ishikawa等(2009)采用亲和免疫层析技术研究大麦HvNRT2.1和HvNAR- 2.3蛋白互作位点, 结果表明HvNAR2.3的中心域与HvNRT2.1的C端(不包含跨膜域)直接互作, 当HvNRT2.1的C端第463位丝氨酸残基(Ser463)突变为丙氨酸(S463A)后, 显著降低了其与HvNAR2.3结合的能力。研究人员提出HvNRT2.1-HvNAR2.3复合物的形成或解离可能影响HvNRT2.1蛋白Ser463的磷酸化或去磷酸化, 从而参与调控NRT2.1的HATS活性及其在质膜上的正确定位。
尽管有些磷酸化位点的功能已经明确, 但对于NRT2.1磷酸化修饰的发生及其作用机制还知之甚少。研究表明, NRT2.1与NAR2蛋白的互作强度可能与NRT2.1蛋白的磷酸化修饰有关。Ishikawa等(2009)采用亲和免疫层析技术研究大麦HvNRT2.1和HvNAR- 2.3蛋白互作位点, 结果表明HvNAR2.3的中心域与HvNRT2.1的C端(不包含跨膜域)直接互作, 当HvNRT2.1的C端第463位丝氨酸残基(Ser463)突变为丙氨酸(S463A)后, 显著降低了其与HvNAR2.3结合的能力。研究人员提出HvNRT2.1-HvNAR2.3复合物的形成或解离可能影响HvNRT2.1蛋白Ser463的磷酸化或去磷酸化, 从而参与调控NRT2.1的HATS活性及其在质膜上的正确定位。
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al., Arabidopsis NIN-like transcription factors have a central role in nitrate signaling 4 2013 ... Transcription factors involved in regulating NRT2
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al., Arabidopsis NIN-like transcription factors have a central role in nitrate signaling 4 2013 ... Transcription factors involved in regulating NRT2
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al., Dual regulation of root hydraulic conductivity and plasma membrane aquaporins by plant nitrate accumulation and high-affinity nitrate transporter NRT2.1 1 2016 ... 此外, 根系中硝酸盐的积累还与根系水分运输密切相关.当AtNRT2.1突变后, 根系的水分运输能力降低30%, 表明AtNRT2.1对根系导水率和质膜水通道蛋白活性具有调控作用.相关数据显示, AtNRT2.1对根系导水率的调控不依赖硝酸根(Li et al., Dissection of the AtNRT2.1:AtNRT2.2 inducible high-affinity nitrate transporter gene cluster 2 2007 ... 迄今为止, 在拟南芥中共鉴定到7个NRT2家族成员, 其中AtNRT2.1、AtNRT2.2、AtNRT2.4和AtNRT2.5属于高亲和硝酸盐转运系统(Orsel et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al., Dual regulation of root hydraulic conductivity and plasma membrane aquaporins by plant nitrate accumulation and high-affinity nitrate transporter NRT2.1 1 2016 ... 此外, 根系中硝酸盐的积累还与根系水分运输密切相关.当AtNRT2.1突变后, 根系的水分运输能力降低30%, 表明AtNRT2.1对根系导水率和质膜水通道蛋白活性具有调控作用.相关数据显示, AtNRT2.1对根系导水率的调控不依赖硝酸根(Li et al., Dissection of the AtNRT2.1:AtNRT2.2 inducible high-affinity nitrate transporter gene cluster 2 2007 ... 迄今为止, 在拟南芥中共鉴定到7个NRT2家族成员, 其中AtNRT2.1、AtNRT2.2、AtNRT2.4和AtNRT2.5属于高亲和硝酸盐转运系统(Orsel et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al., 3 2020 ... 其次, 该领域的一些研究证明, 拟南芥AtNRT2.1蛋白的磷酸化和去磷酸化是调控其HATS活性的重要机制, 同时AtNRT2.1也是目前唯一被报道具有磷酸化修饰的NRT2家族成员(
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al., 3 2020 ... 其次, 该领域的一些研究证明, 拟南芥AtNRT2.1蛋白的磷酸化和去磷酸化是调控其HATS活性的重要机制, 同时AtNRT2.1也是目前唯一被报道具有磷酸化修饰的NRT2家族成员(
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al., Analysis of the NRT2 nitrate transporter family in Arabidopsis. Structure and gene expression 1 2002 ... 迄今为止, 在拟南芥中共鉴定到7个NRT2家族成员, 其中AtNRT2.1、AtNRT2.2、AtNRT2.4和AtNRT2.5属于高亲和硝酸盐转运系统(Orsel et al., Major facilitator superfamily 1 1998 ... NRT2蛋白结构的主要特点: (1) 一般含有500-600个氨基酸残基, 通常具有12个跨膜域(transmembrane helical segments, TMHs).在结构上, 可分为由第1-6个跨膜域组成的N端结构域(TM1-6)和第7-12个跨膜域组成的C端结构域(TM7-12), 2个结构域在序列上缺乏相似性, 但在蛋白结构上相互对称, 它们之间由胞质环相连; (2) 在第2-3个跨膜域之间含有1个特征序列为G-x-x-x-D-x-x-G-x-R的保守基序, 即MFS序列; (3) 在第5个跨膜域中含有保守的NNP序列, 即A-G-W/L-G-N-M/A-G基序; (4) 大多数Type IIIb蛋白在N端和C末端域含有潜在的蛋白激酶C识别基序(S/T-x-R/K) (Pao et al., Dichotomy in the NRT gene families of dicots and grass species 2 2010 ... Summary of identified NRT2 transporters in Arabidopsis thaliana, Oryza sativa and Zea mays
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al., Analysis of the NRT2 nitrate transporter family in Arabidopsis. Structure and gene expression 1 2002 ... 迄今为止, 在拟南芥中共鉴定到7个NRT2家族成员, 其中AtNRT2.1、AtNRT2.2、AtNRT2.4和AtNRT2.5属于高亲和硝酸盐转运系统(Orsel et al., Major facilitator superfamily 1 1998 ... NRT2蛋白结构的主要特点: (1) 一般含有500-600个氨基酸残基, 通常具有12个跨膜域(transmembrane helical segments, TMHs).在结构上, 可分为由第1-6个跨膜域组成的N端结构域(TM1-6)和第7-12个跨膜域组成的C端结构域(TM7-12), 2个结构域在序列上缺乏相似性, 但在蛋白结构上相互对称, 它们之间由胞质环相连; (2) 在第2-3个跨膜域之间含有1个特征序列为G-x-x-x-D-x-x-G-x-R的保守基序, 即MFS序列; (3) 在第5个跨膜域中含有保守的NNP序列, 即A-G-W/L-G-N-M/A-G基序; (4) 大多数Type IIIb蛋白在N端和C末端域含有潜在的蛋白激酶C识别基序(S/T-x-R/K) (Pao et al., Dichotomy in the NRT gene families of dicots and grass species 2 2010 ... Summary of identified NRT2 transporters in Arabidopsis thaliana, Oryza sativa and Zea mays
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
与其它NRT2定位于质膜不同, AtNRT2.7定位于液泡膜, 在干种子中的表达量很高, 并介导种子中NO3-的积累(Chopin et al.,
本文的其它图/表
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
 首页
首页