|
|
|||||||||||||||||||||||||||||||||||||||||||||||||
|
2022年中国植物科学重要研究进展
植物学报
2023, 58 (2):
175-188.
DOI: 10.11983/CBB23061
2022年中国科学家在植物科学主流期刊发表的论文数量相比2021年显著增加, 在防止“多精受精”分子机制, 胞外pH感受器, 叶绿体蛋白运输通道结构, 作物高产、优质、耐逆、抗病及共生固氮机制, 被子植物自交不亲和性的起源和演化机制, 甘蔗和玉蜀黍种质资源演化等方面取得了重要研究进展。其中, “水稻抗高温基因挖掘及调控新机制”入选2022年度中国生命科学十大进展。该文总结了2022年度我国植物科学研究取得的成果, 简要介绍了30项有代表性的重要进展, 梳理了植物科学研究中所使用的实验材料, 以帮助读者了解我国植物科学的发展态势, 进而思考如何更好地开展下阶段研究和服务国家需求。
表1
2020-2022年中国与4个欧美国家的科学家在5种植物科学主流期刊(MP、NP、PC、PP和PJ)的发文量比较(数据来源: Web of Science核心合集)
正文中引用本图/表的段落
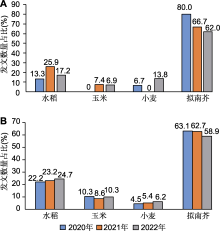
据我刊不完全统计, 2022年中国植物科学家在Science、Cell、Nature及其子刊(Nature Plants、Nature Genetics、Nature Communications)和PNAS期刊上发表的论文(研究类, 如Article和Letter)总数为163篇, 与2021年(161篇)相比略有增长, 而在Cell、Nature和Science三种国际综合性学术期刊发表论文(研究类)数量稍有下降, 为22篇(2021年为24篇); 在Molecular Plant (MP)、Nature Plants (NP)、The Plant Cell (PC)、Plant Physiology (PP)和The Plant Journal (PJ)五种植物科学主流期刊上发表论文742篇, 与2021年(640篇)相比增长16%。近3年的统计数据显示, 2021年中国科学家在上述5种植物科学主流期刊上发表的论文数占这些期刊总载论文量的约44%, 2022年则迅速增长至约50%, 已连续4年(2019-2022年)位居世界第1 (表1) (数据来源: Web of Science核心合集) (检索时间: 2023年3月29日)。
注: 以2个或2个以上物种为材料的文章被重复计数。3种国际综合性学术期刊(Science、Nature和Cell)刊登的以水稻、玉米、小麦和拟南芥为实验材料的文章数量总和分别为15 (2020年)、27 (2021年)和29 (2022年)篇; 5种(MP、NP、PC、PP和PJ)植物科学主流期刊刊登的以水稻、玉米、小麦和拟南芥为实验材料的文章数量总和分别为555 (2020年)、569 (2021年)和681 (2022年)。MP、NP、PC、PP和PJ同
生长素作为第1种被发现的植物激素, 极性运输是其区别于其它植物激素的显著特征之一。转运蛋白在此过程中发挥了关键作用, 其中, 在细胞膜上不对称分布的植物特异性生长素外排蛋白家族PIN尤为重要。但PIN家族蛋白识别和转运生长素的机制并不清楚。最近的研究表明, 在实验室广泛使用的一种生长素极性运输抑制剂, 同时也是农业生产中最早作为除草剂应用的化学小分子NPA可直接结合PIN蛋白(Abas et al., 2021), 但其作用机制未知。孙林峰研究组与国内合作者利用基于放射性同位素的功能检测体系证实了在拟南芥中广泛表达的PIN1蛋白具有转运生长素3-吲哚乙酸(IAA)的活性, 可被蛋白激酶激活并被NPA抑制; 利用哺乳动物表达系统和纳米抗体表达, 他们纯化了拟南芥PIN1蛋白; 利用冷冻电镜单颗粒重构技术成功解析了PIN1单独的(apo)、与底物IAA结合的以及与抑制剂NPA结合的3个高分辨率结构。PIN1的跨膜结构域为保守的NhaA蛋白折叠, 呈现向细胞质侧开放的构象。虽然IAA和NPA在结合方式上具有一定的相似性, 但是NPA以一种更高亲和力的方式结合PIN1, 阐明了NPA的高效抑制作用机制(Yang et al., 2022)。该研究揭示了PIN1识别底物生长素IAA及被NPA抑制的分子机制, 为理解生长素的外排和极性运输提供了结构基础, 也为针对PIN家族蛋白的农业用除草剂和生长调节剂设计开发奠定了理论基础。
全球一直面临耕地资源短缺和粮食产量不足的问题。提高水稻单株产量和缩短生育期可更高效地利用有限的耕地资源, 满足日益增长的粮食需求。而实现水稻增产需提高光合作用效率和氮素利用率, 这些复杂的生物学过程常涉及转录因子的调控。周文彬研究组从候选的118个转录因子中鉴定到1个能同时受光照和低氮诱导的转录因子OsDREB1C。该转录因子可激活光合作用的碳同化、氮吸收转运以及抽穗开花等途径的下游靶基因转录: OsDREB1C能够调控编码Rubisco小亚基的OsRBCS3基因(Suzuki and Ma-kino, 2012), 增强植株的光合能力; 并能提高硝酸盐转运蛋白基因OsNRT1.1B (Zhang et al., 2019)和OsNRT2.4, 及硝酸还原酶基因OsNR2 (Gao et al., 2019)的表达水平, 从而促进氮素的吸收和转运; 还能作用于OsFTL1基因促进早花。将OsDREB1C基因过表达能够显著增加水稻单产、缩短生育期及提高氮素的利用率, 最终实现碳氮的有效分配(Wei et al., 2022)。该研究为未来水稻育种实践提供了重要的靶标基因, 也为通过单基因功能研究优化作物性状提供了范例。
无机化学肥料使用和耕作方式的改变增加了土壤硬度, 高硬度的土壤会限制植物本身产生的乙烯扩散, 导致其在根系周围积聚, 引起根系变短、增粗, 从而响应土壤硬度的增加(Pandey et al., 2021)。然而, 在植物根系感应土壤硬度的过程中, 乙烯转化为植物内部发育信号的分子机制尚不明确。张大兵研究组与国外合作者发现, 乙烯在压实土壤中通过转录因子OsEIL1上调生长素合成基因OsYUC8的表达, 从而增加水稻根系内部生长素的浓度, 并且通过生长素载体OsAUX1将合成的生长素运输至伸长区表皮细胞中, 抑制根伸长。另外, ABA作用于乙烯和生长素的下游, 调控皮层细胞的径向扩张。破坏根尖径向膨胀的ABA缺陷突变体表现出更强的穿透压实土壤的能力, 这打破了径向膨胀有助于根系穿透压实土壤的传统认知(Huang et al., 2022a)。该研究解析了植物将乙烯信号转化为发育信号响应外界土壤硬度的机制, 为未来培育适应不同土壤硬度的作物新品种奠定了重要理论基础。
病原菌入侵植物体, 其相关分子模式(pathogen-associated molecular patterns, PAMPs)首先受到模式识别受体(pattern recognition receptors, PRRs)的识别, 触发植物细胞的PTI (pattern-triggered immunity)反应(Jones and Dangl, 2006)。模式识别受体主要分为受体类激酶(receptor kinases, RKs)和受体类蛋白(receptor-like proteins, RLPs) (Jamieson et al., 2018; Wan et al., 2019)。受体类蛋白的活化和配体识别机制一直未被解析。柴继杰研究组与国内合作者通过解析单独受体(RXEG1)、受体-配体复合体(RXEG1-XEG1)以及受体-配体-共受体复合体(RXEG1- XEG1-BAK1)等处于静息或激活状态的蛋白晶体结构, 发现RXEG1通过其N端和C端的loop结构与XEG1结合, 二者结合后RXEG1的N端和C端结构域发生构象变化, 促进受体RXEG1与共受体BAK1形成异源二聚体。生化实验与功能分析进一步表明, XEG1诱导的RXEG1-BAK1异源二聚体的形成是激活下游抗病信号途径的必要条件。该研究同时发现RXEG1结合部位位于XEG1的酶活区域, RXEG1受体结合后显著抑制XEG1的糖基水解酶活性, 直接抑制病原菌侵染(Sun et al., 2022)。
胞外pH是影响细胞化学和生物学反应的重要因素(Tsai and Schmidt, 2021)。而胞外液碱性化是植物细胞的免疫指标之一。例如, 病原物相关分子模式(PAMP)所触发的免疫反应(PTI)可以引发胞外液pH值升高(Felix et al., 1999; Huffaker et al., 2006)。越来越多的研究表明, 植物细胞可感受外界pH变化, 但其感知pH的机制尚未被揭示。郭红卫研究组与柴继杰研究组合作, 采用遗传、生化和结构解析等手段, 对拟南芥根尖分生区细胞进行了研究, 发现植物细胞利用效应因子RGF1和免疫受体蛋白PEPR响应酸碱环境变化。在胞间液酸性环境下, RGF1的酪氨酸磺酸化修饰位点发生质子化反应, 与受体的RxGG基序产生强烈的氢键相互作用, 增强RGF与下游膜蛋白受体RGI/RGFR的结合能力; 在碱性条件下, RGF1的酪氨酸磺酸化修饰发生去质子化, 导致其结合能力下降, 从而调节植物生长信号的传递。同时发现, 酸性条件下植物免疫受体蛋白PEPR的酸性氨基酸(Asp, Glu)质子化, 破坏了小肽Pep1与PEPR互作, 抑制下游免疫信号途径; 碱性环境下, 该酸性氨基酸则会去质子化, 促进受体与共受体互作, 激活下游免疫反应。互换RGF1和PEPR的胞外受体结构域, 二者对酸碱的响应会发生改变(Liu et al., 2022a)。该研究揭示了“胞外碱化”的免疫反应标志和“酸生长理论”的分子机制, 为“酸碱控制”植物生长、抗逆、抗病育种及农业生产应用奠定了理论基础。《植物学报》发表专文点评, 认为该研究发现了植物细胞质外体pH的感受器, 阐明了质外体碱化的感受机制及植物协调免疫与生长发育的机制, 加深了人们对植物平衡生长与免疫应答生物学反应过程的理解(杨永青和郭岩, 2022)。
抗病育种能有效提高作物的抗病性。R基因在育种中应用广泛, 但R基因赋予的质量抗性易引发病原菌发生突变, 导致短期内作物的抗性丧失(Chen et al., 2014)。研究发现, 由突变感病基因所赋予植物的抗性更具有持久性和广谱性(Pavan et al., 2010; Lang-ner et al., 2018)。近期, 王晓杰研究组与国内合作者克隆了1个小麦条锈病感病基因TaPsIPK1, 该基因突变后可赋予小麦对多种条锈菌的广谱抗性。他们以TaPsIPK1为诱饵与条锈菌蛋白库进行酵母双杂交, 鉴定到其互作效应因子PsSpg1; 进一步生化实验证明, PsSpg1可以通过激活TaPsIPK1自磷酸化促进植物感病, 并且磷酸化后TaPsIPK1由质膜转移至细胞核; 免疫共沉淀联合质谱分析, 发现转录因子TaCBF1可以与TaPsIPK1互作, 后者通过抑制TaCBF1介导的下游抗病基因的转录活性而促进植株的感病性增强(Wang et al., 2022b)。该研究鉴定出小麦受锈菌效应因子操控的感病基因, 为条锈病和叶锈病持久且广谱抗性育种提供了极具价值的种质资源。《植物学报》发表专文点评, 认为该研究揭示了由PsSpg1-TaPsIPK1-TaCBF1d在小麦条锈病S基因中触发的新的磷酸化转录调控机制, 为通过作物遗传修饰培育持久抗性品种提供了新策略(孔令让, 2022)。
Mlo (Mildew resistance locus O)是最早在大麦中克隆的感病基因, 因其序列和功能的保守性, 该基因突变能赋予多种植物对白粉病的广谱抗性(Büschges et al., 1997; Schulze-Lefert and Vogel, 2000)。前期, 高彩霞研究组与国内合作者通过基因编辑创制了六倍体小麦Tamlo突变体, 该突变体具有白粉病抗性, 但株高和产量明显下降(Wang et al., 2014)。近期, 他们利用TALEN对小麦品种Bobwhite背景下A、B、D基因组的MLO1基因进行沉默, 创制出新型mlo突变体Tamlo-R32, 该突变体不仅保留了白粉病广谱抗性, 而且发育表型和产量均正常。通过遗传分析和基因组重测序, 发现该突变体除在TaMLO-A1以及TaMLO-D1基因处发生编辑外, 还在TaMLO-B1基因座附近发生了约304 kb的大片段删除, 进而造成该部分染色质形态发生改变, 表观调控TaTMT3B基因的表达, 该基因超表达可恢复TaMLO的产量和发育表型。此外, 与mlo相同的是TMT3B的功能也具有保守性, 在拟南芥中超表达TMT3B可以缓解拟南芥mlo突变体的早衰表型。进一步, 他们通过传统杂交育种和CRISPR/Cas9基因编辑技术在多个小麦主栽品种中实现了Tamlo-R32类突变(Li et al., 2022a)。该研究证实了利用基因编辑技术可对多倍体作物复杂的多性状实现精准调控, 为作物抗病育种研究提供了新的理论视角。
林鸿宣研究组还定位并克隆了编码G蛋白γ亚基的基因TT2。热带粳稻来源的TT2存在1个SNP, 该SNP使其编码1个提前终止形式的蛋白, NIL-TT2HPS32植株具有较强的耐热性, 且成熟期的单株产量显著提高; 而在高温敏感的温带粳稻中, 该SNP占比较低。在高温敏感的对照株系中, 蜡质相关调控基因受热诱导显著下调, 而在NIL-TT2HPS32株系中有一部分蜡质基因(包括1个正向调控蜡质合成的重要转录因子OsWR2基因)不响应热并且表达稳定。敲除NIL-TT2HPS32株系中OsWR2使其耐高温表型消失, 表明在高温胁迫下维持正常的蜡质含量对水稻的耐热性至关重要。进一步研究发现水稻钙调素结合转录因子(CAMTA)家族成员SCT1可直接结合OsWR2的启动子, 影响OsWR2的表达, 负调控水稻的耐热性。研究还发现高温提高细胞内的Ca2+浓度, 诱导SCT1与钙调素相互作用, 抑制OsWR2的表达(Kan et al., 2022)。该研究鉴定到水稻耐热性负调控因子TT2, 并揭示了TT2-SCT1-OsWR2模块调控水稻耐热性的分子机制。
玉米起源于热带, 低温胁迫严重威胁着玉米生产。然而, 目前人们对玉米响应低温的分子机制研究尚处于起步阶段, 可利用的优势基因资源匮乏。杨淑华研究组对245份自交系材料进行了常温和低温处理下的非靶标代谢组学分析及mGWAS分析, 发现转录因子ZmICE1参与低温诱导的代谢调控过程。过表达ZmICE1可以显著增强玉米萌发期以及苗期的耐冷性, 表明ZmICE1是玉米耐冷性的正调控因子。利用RNA-seq及ChIP-seq鉴定了802个ZmICE1的直接靶基因, 除ZmDREB1s和ZmERFs等冷响应基因外, 超过半数的靶基因参与各种代谢过程。其中一类ZmASs (ASPARAGINE SYNTHETASES)基因编码Glu-Asn合成关键酶, 在氮代谢中发挥重要作用。通过候选基因关联分析, 发现ZmICE1启动子区SNP-465位点变异影响了ZmMYB39转录因子与ZmICE1启动子的结合, 从而影响ZmICE1的转录水平。耐冷单倍型中, ZmMYB39与ZmICE1启动子区的结合能力增强, ZmICE1转录水平增高。 ZmICE1蛋白一方面促进冷响应基因(如ZmDREB1s)的表达; 另一方面通过抑制ZmAS的表达降低Glu/Asn的生物合成, 从而减少由Glu引起的线粒体活性氧(mtROS)的产生, 解除mtROS对DREB1s的抑制(Jiang et al., 2022)。该研究揭示了转录因子ZmICE1调控玉米耐低温胁迫与氨基酸代谢的分子机制, 为作物性状改良提供了新思路。
本文的其它图/表
|
|||||||||||||||||||||||||||||||||||||||||||||||||
 首页
首页