
|
|
||
|
白菜类作物抗TuMV研究进展
植物学报
2023, 58 (2):
335-346.
DOI: 10.11983/CBB22061
白菜类作物主要包括大白菜(Brassica rapa subsp. pekinensis)、不结球白菜(B. rapa subsp. chinensis)和芜菁(B. rapa subsp. rapa)等, 可用作蔬菜、油料及饲料。白菜类作物在生产过程中常受到病毒病的危害, 其中芜菁花叶病毒(TuMV)是主要病原物。近年来, 在TuMV抗性基因的遗传分析、定位克隆及分子机制方面取得了一定研究进展, 为培育优质抗病新品种奠定了基础。该文对国内外关于白菜类作物抗TuMV的遗传分析、抗性基因与分子标记、生物信息学与组学分析以及基因工程技术在抗TuMV研究中的应用进行综述, 以期为白菜类作物抗TuMV研究和技术利用提供科学依据。 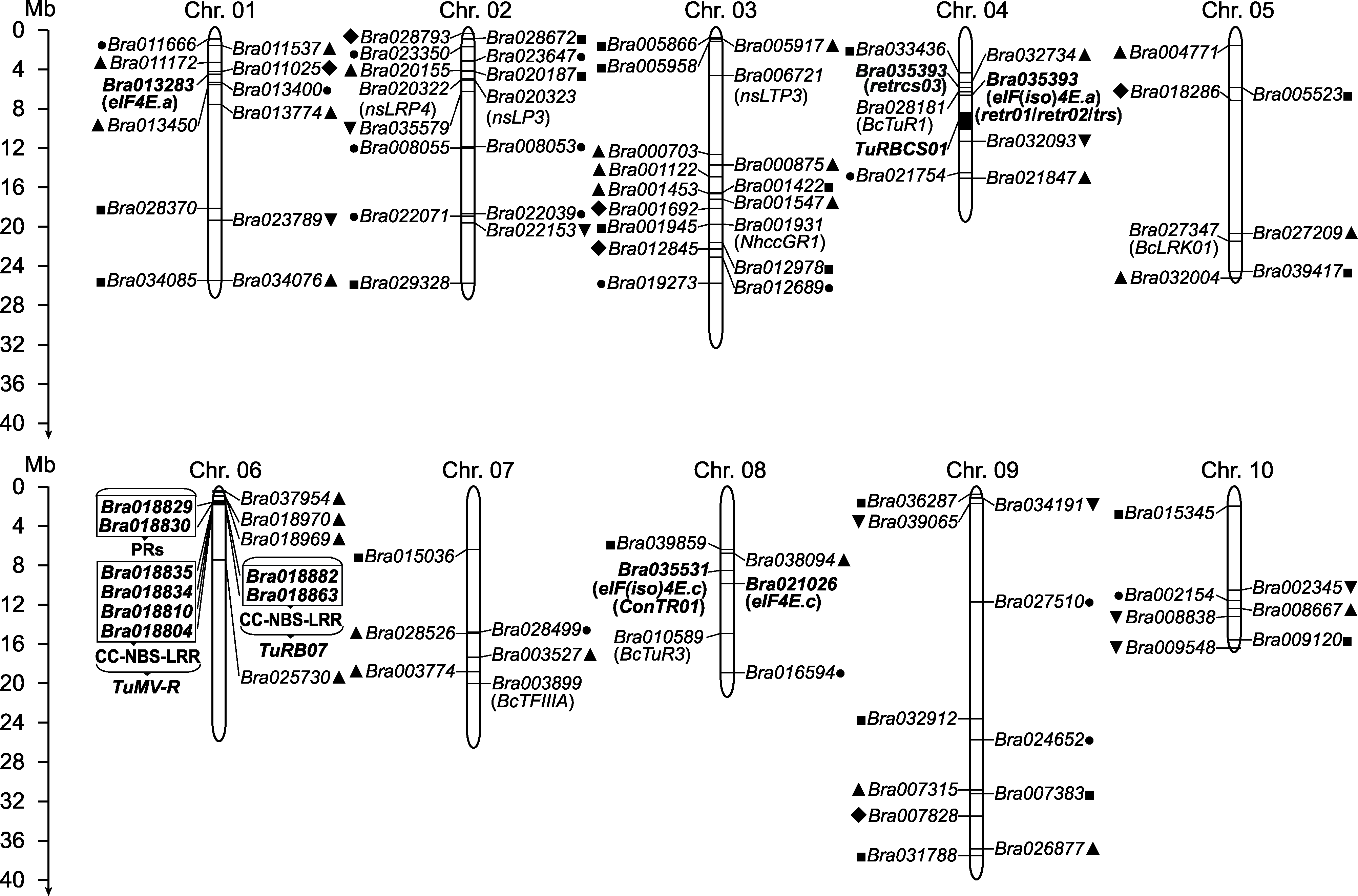 View image in article
图1
TuMV抗性基因及位点在白菜染色体上的分布
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点。黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因。实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因。为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置。本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
正文中引用本图/表的段落
芸薹属中发现的TuMV抗性基因主要集中在白菜类作物的A基因组中, 鉴定到的抗性基因既有显性也有隐性基因。显性抗病基因多属于R基因, R基因通过激活超敏反应(hypersensitive response, HR)和系统获得性抗性(systemic acquired resistance, SAR)抵抗病毒。目前, 在大白菜中发现的显性基因(或标记)有ConTR01、TuRB01b、Rnt1-1、TuRB07、Tu1-Tu4、TuR1-TuR4和Tu1-Tu3等(表1; 图1)。Rusholme等(2007)利用RFLP标记鉴定到大白菜抗TuMV基因ConTR01 (conditional TuMV resistance 01), 该基因位于R08连锁群上, 后证实BraA.eIF(iso)4E.c是其唯一候选基因(Nellist et al., 2014)。Lydiate等(2014)在大白菜中鉴定到TuRB01b (TuMV RESISTANCE IN BRASSICA 01b), 其位于A06染色体上, 研究发现它与油菜中的TuRB01同源或互为等位基因, 经分析认为油菜从白菜的A基因组中获得了该基因。
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点。黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因。实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因。为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置。本图基于Brara_Chiifu_V1.5基因组(Wang et al., 2011b)使用MG2C v2.1软件(Chao et al., 2021)绘制。
Fujiwara等(2011)筛选到的Rnt1-1 (resistance and necrosis to TuMV 1)位点位于R06连锁群, 但与TuRB01b不同。TuMV自然突变体TuMV-UK1 (UK1m)的CI突变(V1827E)可以打破Rnt1-1的抗性, 后发现Rnt1-1可以提高白菜内源抗坏血酸(ascorbic acid, AS)和脱氢抗坏血酸(dehydroascorbic acid, DHA)的含量, 从而增强植物的抗病能力(Fujiwara et al., 2016)。后续又发现了多个显性抗病位点。例如, Chung等(2014)定位到了TuMV-R位点, 其中包含4个CC-NBS-LRRs (coiled-coil nucleotide-binding site leucine-rich repeats)基因和1个病程相关基因PR (pathogenesis-related gene)。Jin等(2014)鉴定到的TuRB07位点包含2个CC-NBS-LRRs基因。目前, 通过分子标记筛选到的显性抗病基因或分子标记主要集中在A06染色体上, 且成簇分布, 说明此区域很可能存在多个抗TuMV的CC-NBS-LRRs和PRs基因(图1)。Li等(2015)利用SSR (simple sequence repeats)标记在大白菜中鉴定到1.98 Mb的TuRBCS01显性位点。除了上述抗性基因外, 还得到多个与抗TuMV相关的QTLs (quantitative trait loci)。Zhang等(2008)通过对100株DH系接种TuMV-C4, 得到4个QTLs, 其中2个QTLs (Tu1和Tu2)与苗期抗性相关, 2个QTLs (Tu3和Tu4)与成株期抗性相关。张俊华等(2008)利用A52-2高抗大白菜自交系对TuMV-C3进行分析, 得到4个QTLs (TuR1-TuR4)。张晓伟等(2009)利用DH系对TuMV-C4抗性进行分析, 得到3个抗病相关QTLs (Tu1-Tu3)。
Liu等(2017)鉴定到eIF(iso)4E.a和eIF(iso)4E.c双位点功能缺失突变体自交系材料He102, 可作为遗传背景更丰富的抗性材料。Kim等(2013)在大白菜中发现了抗病基因trs (TuMV resistance discovered at Seoul National University), 该基因位于第4号染色体, 与retr02紧密连锁且位点存在重叠, 因此trs很可能与retr01和retr02为同一基因。目前筛选到的隐性抗TuMV基因主要是eIF(iso)4Es或与其有紧密连锁关系的基因(图1)。满卫萍(2020)对以大白菜TuMV抗病材料73和感病材料06-247为亲本构建的F2和BC1群体进行分析, 得到1个隐性抗病基因retrcs03, 候选基因是Bra032679, 编码γ-干扰素诱导蛋白。
在不结球白菜中也鉴定到多个抗TuMV基因和标记。马景蕃等(2009)根据拟南芥(Arabidopsis thaliana)等物种的同源基因, 通过RACE技术从不结球白菜中克隆到BcTuR1和BcTuR3, 它们都受TuMV诱导高表达, 且BcTuR3属于TIR-NBS-LRR家族蛋白。在不结球白菜中, Wang等(2011a)通过AFLP标记定位到抗TuMV-C5的TuRBCH01基因。彭海涛等(2012a)和李彦肖等(2012)都通过cDNA-AFLP技术, 分别从感染TuMV的不结球白菜中克隆到BcLRK01和NhccGR1, 并证明这2个基因可能是不结球白菜的病程相关基因。但BcTuR1、BcTuR3、BcLRK01和NhccGR1尚未被确定是抗性基因, 也未分析其易感等位基因, 因此属于抗性候选基因, 尚待进一步验证。根据图1中抗性基因的染色体分布, 后续应特别关注定位于A04、A06和A08染色体上的基因。
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al., 大白菜TuMV抗性的主基因+多基因混合遗传分析 1 2012 ... 白菜抗TuMV的遗传规律非常复杂, 国内外相关研究很多, 但结论不一.早在1980年, Provvidenti ( 大白菜抗TuMV-C3的QTL分析 1 2009 ... 白菜抗TuMV的遗传规律非常复杂, 国内外相关研究很多, 但结论不一.早在1980年, Provvidenti ( 双价反义载体构建及对大白菜的遗传转化 1 2020a ... 目前, 研究人员已获得多个抗性相关的基因, 多种生物技术也应用于白菜品质改良(刘栓桃等, 无选择标记转基因大白菜的检测 1 2020b ... 目前, 研究人员已获得多个抗性相关的基因, 多种生物技术也应用于白菜品质改良(刘栓桃等, 不结球白菜抗芜菁花叶病毒基因分子标记与遗传定位 1 2010 ... 目前, 对不结球白菜的遗传模型研究不多, 但同样存在多样性.曹光亮和曹寿椿( 大白菜BrCNGC全基因组鉴定及其表达分析 1 2018 ... 除了通过分子标记鉴定TuMV抗性相关基因外, 研究人员还通过生物信息学手段预测或进行多组学分析得到大量候选基因.基因家族是结构和功能上具有相似性的一组基因, 筛选鉴定与病毒相关的基因家族是快速高效获得候选基因的方法.Lv等( 大白菜、萝卜芜菁花叶病毒系统进化及CP序列分析 1 2018 ... 随着科技的进步, 分子生物学技术的应用使病毒株系鉴定更加精确和灵敏.研究人员使用抗体从血清学角度对TuMV进行了分类鉴定.Jenner等( 1 2000 ... 白菜抗TuMV的遗传规律非常复杂, 国内外相关研究很多, 但结论不一.早在1980年, Provvidenti ( 白菜病毒诱导基因沉默技术体系的建立 1 2012 ... 综上, 白菜类作物抗TuMV相关研究取得了较大进展, 已发现多个抗性基因, 为今后分子育种和抗病毒品种的选育奠定了良好的基础.同时还可为芸薹属B、C基因组的抗病品种选育提供基因资源, 并为抗病毒分子育种提供重要参考.下一步可通过远缘杂交、体细胞融合或基因工程技术将抗性基因导入B、C基因组, 从而创制新的抗病材料.目前, 芸薹属作物特别是A基因组作物的转基因技术尚不成熟.作为替代病毒诱导的基因沉默技术已在白菜类作物中广泛应用, 可实现在不转基因的情况下快速对筛选到的候选基因进行功能验证(杨学东等, TuMV-Nib反义基因对大白菜的遗传转化研究 1 2006 ... 目前, 研究人员已获得多个抗性相关的基因, 多种生物技术也应用于白菜品质改良(刘栓桃等, 大白菜抗芜菁花叶病毒的QTL分析 3 2008 ... 白菜抗TuMV的遗传规律非常复杂, 国内外相关研究很多, 但结论不一.早在1980年, Provvidenti (
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al., Correlation analysis of expression profile and quantitative iTRAQ-LC-MS/MS proteomics reveals resistance mechanism against TuMV in Chinese cabbage (Brassica rapa ssp 1 2020 ... 与基因家族分析相比, 组学分析可以获得更高通量且更全面的数据.Wang等( Cloning and characterization of the BcTuR3 gene related to resistance to turnip mosaic virus (TuMV) from non-heading Chinese cabbage 1 2010 ... The TuMV resistance genes and makers in Brassica rapa
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al., Ultrastructure of cylindrical inclusions induced by viruses of the potato Y group as visualized by freeze-etching 1 1974 ... 1963年, Yoshii ( Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization 1 1935 ... 白菜(Brassica rapa)属于十字花科(Cruciferae)芸薹属(Brassica), 染色体组为AA, 倍性2n=2x=20.其原产中国, 栽培历史悠久, 形态多样, 遗传资源丰富.白菜类作物是我国分布最广且种植面积最大的一类园艺作物, 主要包括大白菜(B. rapa subsp. pekinensis)、不结球白菜(B. rapa subsp. chinensis)和芜菁(B. rapa subsp. rapa)等亚种.白菜类作物不仅可用作蔬菜、油料及饲料, 还为甘蓝型油菜(B. napus, 染色体组为AACC)和芥菜(B. juncea, 染色体组为AABB)提供了A染色体组(Nagaharu, Turnip mosaic virus, a virus for all seasons 2 2022 ... TuMV主要通过蚜虫以非持久性的方式进行传播(Nellist et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al., Genome-wide identification of turnip mosaic virus-responsive microRNAs in non-heading Chinese cabbage by high-throughput sequencing 1 2015 ... 与基因家族分析相比, 组学分析可以获得更高通量且更全面的数据.Wang等( The SNARE protein Syp71 is essential for turnip mo- saic virus infection by mediating fusion of virus-induced vesicles with chloroplasts 1 2013 ... 除了通过分子标记鉴定TuMV抗性相关基因外, 研究人员还通过生物信息学手段预测或进行多组学分析得到大量候选基因.基因家族是结构和功能上具有相似性的一组基因, 筛选鉴定与病毒相关的基因家族是快速高效获得候选基因的方法.Lv等( Inheritance of resistance to turnip mosaic virus in Chinese cabbage 1 1993 ... 白菜抗TuMV的遗传规律非常复杂, 国内外相关研究很多, 但结论不一.早在1980年, Provvidenti ( On the strain distribution of turnip mosaic virus 1 1963 ... 1963年, Yoshii ( The methylation pattern of DNA and complex correlations with gene expressions during TuMV infection in Chinese cabbage 1 2019 ... 与基因家族分析相比, 组学分析可以获得更高通量且更全面的数据.Wang等( Efficient virus-induced gene silencing in Brassica rapa using a turnip yellow mosaic virus vector 1 2018 ... 综上, 白菜类作物抗TuMV相关研究取得了较大进展, 已发现多个抗性基因, 为今后分子育种和抗病毒品种的选育奠定了良好的基础.同时还可为芸薹属B、C基因组的抗病品种选育提供基因资源, 并为抗病毒分子育种提供重要参考.下一步可通过远缘杂交、体细胞融合或基因工程技术将抗性基因导入B、C基因组, 从而创制新的抗病材料.目前, 芸薹属作物特别是A基因组作物的转基因技术尚不成熟.作为替代病毒诱导的基因沉默技术已在白菜类作物中广泛应用, 可实现在不转基因的情况下快速对筛选到的候选基因进行功能验证(杨学东等, Genome-wide identification, classification, and expression analysis of SNARE genes in Chinese cabbage (Brassica rapa ssp. pekinensis) infected by turnip mosaic virus 1 2018 ... 除了通过分子标记鉴定TuMV抗性相关基因外, 研究人员还通过生物信息学手段预测或进行多组学分析得到大量候选基因.基因家族是结构和功能上具有相似性的一组基因, 筛选鉴定与病毒相关的基因家族是快速高效获得候选基因的方法.Lv等( Quantitative trait loci analysis for resistance against turnip mosaic virus based on a doubled-haploid population in Chinese cabbage 2 2008 ... The TuMV resistance genes and makers in Brassica rapa
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al.,
本文的其它图/表
|
 首页
首页{kind=link}