白菜类作物抗TuMV研究进展
吕善武, 张昌伟, 侯喜林, 邓书林
植物学报
2023, 58 ( 2):
335-346.
DOI: 10.11983/CBB22061
白菜类作物主要包括大白菜(Brassica rapa subsp. pekinensis)、不结球白菜(B. rapa subsp. chinensis)和芜菁(B. rapa subsp. rapa)等, 可用作蔬菜、油料及饲料。白菜类作物在生产过程中常受到病毒病的危害, 其中芜菁花叶病毒(TuMV)是主要病原物。近年来, 在TuMV抗性基因的遗传分析、定位克隆及分子机制方面取得了一定研究进展, 为培育优质抗病新品种奠定了基础。该文对国内外关于白菜类作物抗TuMV的遗传分析、抗性基因与分子标记、生物信息学与组学分析以及基因工程技术在抗TuMV研究中的应用进行综述, 以期为白菜类作物抗TuMV研究和技术利用提供科学依据。
| 抗性基因 | 植物材料 | 抗性品种 | 遗传/克隆情况 | TuMV株系 | 参考文献 | | 显性基因/标记 | | | | | ConTR01
| 大白菜
| RLR22
| 定位于R04, 候选基因可能是eIF(iso)4E
| 1、3、4、7、
8、9和12 | Rusholme et al., 2007
| Tu1-Tu4
| 大白菜
| 91-112
| 4个QTLs, Tu1位于LG5连锁群, Tu2位于LG10连锁群, Tu3位于LG3连锁群, Tu4位于LG4连锁群 | C4
| Zhang et al., 2008
| TuR1-TuR4
| 大白菜
| A52-2
| TuR1和TuR2位于LG3连锁群, TuR3和TuR4位于LG7连锁群 | C3
| 张俊华等, 2008
| Tu1-Tu3
| 大白菜
| Y195-93
| Tu1位于R03连锁群, Tu2位于R04连锁群, Tu3位于R06连锁群 | C4
| 张晓伟等, 2009
| Rnt1-1
| 大白菜
| AS9
| 定位于R06连锁群, 与内源抗坏血酸和脱氢抗坏血酸合成相关 | UK1
| Fujiwara et al., 2011, 2016 | | TuRBCH01 | 不结球白菜 | Q048 | 定位于R06连锁群 | C5 | Wang et al., 2011 | TuRB01b
| 大白菜
| TD34-S1
| 定位于A06染色体, 与油菜TuRB01位于同一位点或互为等位基因 | UK1
| Lydiate et al., 2014
| TuRB07
| 大白菜
| VC1/VC40
| 定位于A06染色体, 候选基因是Bra018863, 属于CC-NBS-LRR基因 | C4
| Jin et al., 2014
| TuMV-R
| 大白菜
| VC40
| 定位于A06染色体的0.34 Mb区域, 有6个候选基因, 4个CC-NBS-LRRs基因, 2个病程相关基因 | C4
| Chung et al., 2014
| | TuRBCS01 | 大白菜 | 8407 | 定位于A04染色体的1.98 Mb区域 | C4 | Li et al., 2015 | | 隐性基因/标记 | | | | | retr01
| 大白菜
| RLR22
| 定位于R04连锁群, 候选基因可能是eIF(iso)4E
| 1、3、4、7、
8、9和12 | Rusholme et al., 2007
| retr02
| 大白菜
| BP8407
| 定位于A04染色体, 候选基因是Bra035393, 编码eIF(iso)4E蛋白 | C4
| Qian et al., 2013
| trs
| 大白菜
| SB18/SB22
| 定位于A04染色体, 可能与retr02紧密连锁或是其等位基因 | CHN2、CHN3、CHN4和CHN5 | Kim et al., 2013
| retrcs03
| 大白菜
| 73
| 定位于A04染色体, 候选基因是Bra032679, 编码γ-干扰素诱导蛋白 | C4
| 满卫萍, 2020
| | 其它候选基因 | | | | | | BcTuR1 | 不结球白菜 | 短白梗 | 抗性候选基因, 受TuMV诱导高表达 | C4 | 马景蕃等, 2009 | BcTuR3
| 不结球白菜
| 短白梗
| 抗性候选基因, 受TuMV诱导高表达, 属于TIR- NB-LRR基因 | C4
| Ma et al., 2010
| BcLRK01
| 不结球白菜
| 泰三抗
| 抗性候选基因, 受TuMV诱导高表达, 属于LRR- RLKs基因 | C4
| 彭海涛等, 2012b
| NhccGR1
| 不结球白菜
| 泰三抗
| 抗性候选基因, 受TuMV诱导高表达, 属于谷胱甘肽还原酶基因 | C4
| 李彦肖等, 2012
| | BcTFIIIA | 不结球白菜 | NHCC001 | TuMV抗性基因, 编码锌指转录因子 | C4 | Zhang et al., 2022b |
View table in article
表1
白菜抗TuMV基因及标记
正文中引用本图/表的段落
白菜抗TuMV的遗传规律非常复杂, 国内外相关研究很多, 但结论不一。早在1980年, Provvidenti (1980)使用多个大白菜品种进行了TuMV抗性分析, 发现大白菜对TuMV具有株系特异抗性, 其抗性基因为显性遗传。Leung和Williams (1983)研究表明, TuMV-C1毒株也受显性基因控制。1984年, 我国学者钮心恪(1984)分析了抗、感亲本杂交后代F1、F2和BC1, 表明大白菜对TuMV的抗性表现为显性遗传, 由2个相互独立的基因控制。后续又有多项研究得到了相似的结论(Suh et al., 1995; 阎瑾琦, 2000; Jin et al., 2014)。但Yoon等(1993)使用高抗材料0-2 (高抗TuMV-C1、C2、C3、C4和C5毒株)和3个感病材料及其F1、F2和BC1进行遗传分析, 发现了2个抗TuMV-C4和TuMV-C5毒株的隐性基因。后续也有研究得出大白菜对TuMV的抗性表现为隐性遗传的结论(韩和平, 2003; Rusholme et al., 2007; Li et al., 2011; 满卫萍, 2020)。与以上2种模式不同, 鹿英杰等(1988)研究表明, 大白菜对TuMV的抗性符合加性-显性效应模型, 属于数量遗传。张俊华等(2008)和屈淑平等(2009)分别用高抗TuMV-C3株系的自交系A52-2与感病材料进行分析, 均得到了相同的结论, 即大白菜对TuMV- C3毒株的抗性为不完全显性。张明等(2011)用高抗材料A52-2进行分析, 发现大白菜对TuMV-C4的抗性也是不完全显性, 至少受5对显性基因控制。钱伟等(2012)使用ELISA方法对接种TuMV的大白菜进行鉴定, 分析了P1、P2、F1和F2四世代遗传模型, 表明TuMV-C4抗性符合2对主基因+多基因的遗传模式, 支持数量性状基因控制TuMV抗性的观点。李国亮等(2019)用高抗TuMV的89B和高感的强势为亲本构建 F2群体并进行分析, 结果也支持数量遗传模型。
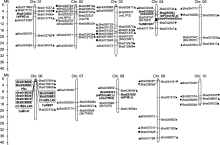
芸薹属中发现的TuMV抗性基因主要集中在白菜类作物的A基因组中, 鉴定到的抗性基因既有显性也有隐性基因。显性抗病基因多属于R基因, R基因通过激活超敏反应(hypersensitive response, HR)和系统获得性抗性(systemic acquired resistance, SAR)抵抗病毒。目前, 在大白菜中发现的显性基因(或标记)有ConTR01、TuRB01b、Rnt1-1、TuRB07、Tu1-Tu4、TuR1-TuR4和Tu1-Tu3等(表1; 图1)。Rusholme等(2007)利用RFLP标记鉴定到大白菜抗TuMV基因ConTR01 (conditional TuMV resistance 01), 该基因位于R08连锁群上, 后证实BraA.eIF(iso)4E.c是其唯一候选基因(Nellist et al., 2014)。Lydiate等(2014)在大白菜中鉴定到TuRB01b (TuMV RESISTANCE IN BRASSICA 01b), 其位于A06染色体上, 研究发现它与油菜中的TuRB01同源或互为等位基因, 经分析认为油菜从白菜的A基因组中获得了该基因。
吕善武( 2019). TuMV侵染下的大白菜表达谱与蛋白组联合分析和TNL & SNARE基因分析验证. 博士论文. 南京: 南京农业大学. pp. 1- 103. [本文引用: 1]
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al., 2011b)使用MG2C v2.1软件(Chao et al., 2021)绘制. ... 大白菜TuMV抗性的主基因+多基因混合遗传分析 1 2012 ... 白菜抗TuMV的遗传规律非常复杂, 国内外相关研究很多, 但结论不一.早在1980年, Provvidenti (1980)使用多个大白菜品种进行了TuMV抗性分析, 发现大白菜对TuMV具有株系特异抗性, 其抗性基因为显性遗传.Leung和Williams (1983)研究表明, TuMV-C1毒株也受显性基因控制.1984年, 我国学者钮心恪(1984)分析了抗、感亲本杂交后代F1、F2和BC1, 表明大白菜对TuMV的抗性表现为显性遗传, 由2个相互独立的基因控制.后续又有多项研究得到了相似的结论(Suh et al., 1995; 阎瑾琦, 2000; Jin et al., 2014).但Yoon等(1993)使用高抗材料0-2 (高抗TuMV-C1、C2、C3、C4和C5毒株)和3个感病材料及其F1、F2和BC1进行遗传分析, 发现了2个抗TuMV-C4和TuMV-C5毒株的隐性基因.后续也有研究得出大白菜对TuMV的抗性表现为隐性遗传的结论(韩和平, 2003; Rusholme et al., 2007; Li et al., 2011; 满卫萍, 2020).与以上2种模式不同, 鹿英杰等(1988)研究表明, 大白菜对TuMV的抗性符合加性-显性效应模型, 属于数量遗传.张俊华等(2008)和屈淑平等(2009)分别用高抗TuMV-C3株系的自交系A52-2与感病材料进行分析, 均得到了相同的结论, 即大白菜对TuMV- C3毒株的抗性为不完全显性.张明等(2011)用高抗材料A52-2进行分析, 发现大白菜对TuMV-C4的抗性也是不完全显性, 至少受5对显性基因控制.钱伟等(2012)使用ELISA方法对接种TuMV的大白菜进行鉴定, 分析了P1、P2、F1和F2四世代遗传模型, 表明TuMV-C4抗性符合2对主基因+多基因的遗传模式, 支持数量性状基因控制TuMV抗性的观点.李国亮等(2019)用高抗TuMV的89B和高感的强势为亲本构建 F2群体并进行分析, 结果也支持数量遗传模型. ... 大白菜抗TuMV-C3的QTL分析 1 2009 ... 白菜抗TuMV的遗传规律非常复杂, 国内外相关研究很多, 但结论不一.早在1980年, Provvidenti (1980)使用多个大白菜品种进行了TuMV抗性分析, 发现大白菜对TuMV具有株系特异抗性, 其抗性基因为显性遗传.Leung和Williams (1983)研究表明, TuMV-C1毒株也受显性基因控制.1984年, 我国学者钮心恪(1984)分析了抗、感亲本杂交后代F1、F2和BC1, 表明大白菜对TuMV的抗性表现为显性遗传, 由2个相互独立的基因控制.后续又有多项研究得到了相似的结论(Suh et al., 1995; 阎瑾琦, 2000; Jin et al., 2014).但Yoon等(1993)使用高抗材料0-2 (高抗TuMV-C1、C2、C3、C4和C5毒株)和3个感病材料及其F1、F2和BC1进行遗传分析, 发现了2个抗TuMV-C4和TuMV-C5毒株的隐性基因.后续也有研究得出大白菜对TuMV的抗性表现为隐性遗传的结论(韩和平, 2003; Rusholme et al., 2007; Li et al., 2011; 满卫萍, 2020).与以上2种模式不同, 鹿英杰等(1988)研究表明, 大白菜对TuMV的抗性符合加性-显性效应模型, 属于数量遗传.张俊华等(2008)和屈淑平等(2009)分别用高抗TuMV-C3株系的自交系A52-2与感病材料进行分析, 均得到了相同的结论, 即大白菜对TuMV- C3毒株的抗性为不完全显性.张明等(2011)用高抗材料A52-2进行分析, 发现大白菜对TuMV-C4的抗性也是不完全显性, 至少受5对显性基因控制.钱伟等(2012)使用ELISA方法对接种TuMV的大白菜进行鉴定, 分析了P1、P2、F1和F2四世代遗传模型, 表明TuMV-C4抗性符合2对主基因+多基因的遗传模式, 支持数量性状基因控制TuMV抗性的观点.李国亮等(2019)用高抗TuMV的89B和高感的强势为亲本构建 F2群体并进行分析, 结果也支持数量遗传模型. ... 双价反义载体构建及对大白菜的遗传转化 1 2020a ... 目前, 研究人员已获得多个抗性相关的基因, 多种生物技术也应用于白菜品质改良(刘栓桃等, 2004), 将这些基因转入白菜或将其突变, 可增强TuMV抗性, 从而提高白菜的产量和品质, 并降低农药使用量.鉴于前期研究发现eIF(iso)4E的突变体对TuMV具有广谱抗性, Kim等(2014)将eIF(iso)4E (W95L、W95L/ K150E)转入大白菜感病品种, 发现可提高TuMV抗性.Nellist等(2014)发现retr01和retr02均为BraA.eIF (iso)4E.a的插入突变体, 突变后基因不能正常剪接, 因此不能被TuMV利用, 将其转入拟南芥eIF(iso)4E突变体后得到抗性植株.除以上直接转入抗性基因外, 还可以将TuMV的编码基因设计成RNAi靶标, 然后转入白菜类作物, 从而提高抗病性.例如, 朱常香等(2001)将TuMV的CP基因转入大白菜, 明显增强其抗病能力.于占东等(2006)和佟容等(2020b)将TuMV的NIb反义片段转入大白菜, 得到多个抗TuMV-C4植株.为了获得既抗TuMV又耐抽薹的大白菜, 佟容等(2020a)利用TuMV-Nib和FLY基因构建了双价反义表达载体并转化大白菜, 最终获得7株同时具有抗病毒和耐抽薹特性的植株. ... 无选择标记转基因大白菜的检测 1 2020b ... 目前, 研究人员已获得多个抗性相关的基因, 多种生物技术也应用于白菜品质改良(刘栓桃等, 2004), 将这些基因转入白菜或将其突变, 可增强TuMV抗性, 从而提高白菜的产量和品质, 并降低农药使用量.鉴于前期研究发现eIF(iso)4E的突变体对TuMV具有广谱抗性, Kim等(2014)将eIF(iso)4E (W95L、W95L/ K150E)转入大白菜感病品种, 发现可提高TuMV抗性.Nellist等(2014)发现retr01和retr02均为BraA.eIF (iso)4E.a的插入突变体, 突变后基因不能正常剪接, 因此不能被TuMV利用, 将其转入拟南芥eIF(iso)4E突变体后得到抗性植株.除以上直接转入抗性基因外, 还可以将TuMV的编码基因设计成RNAi靶标, 然后转入白菜类作物, 从而提高抗病性.例如, 朱常香等(2001)将TuMV的CP基因转入大白菜, 明显增强其抗病能力.于占东等(2006)和佟容等(2020b)将TuMV的NIb反义片段转入大白菜, 得到多个抗TuMV-C4植株.为了获得既抗TuMV又耐抽薹的大白菜, 佟容等(2020a)利用TuMV-Nib和FLY基因构建了双价反义表达载体并转化大白菜, 最终获得7株同时具有抗病毒和耐抽薹特性的植株. ... 不结球白菜抗芜菁花叶病毒基因分子标记与遗传定位 1 2010 ... 目前, 对不结球白菜的遗传模型研究不多, 但同样存在多样性.曹光亮和曹寿椿(1995)通过双列杂交遗传分析, 发现不结球白菜对TuMV的抗性为不完全显性数量遗传.刘琳(2008)通过5世代联合分析, 发现不结球白菜对TuMV的抗性符合1对加性-显性主基因+ 加性-显性多基因模型.还有研究得出的遗传模型为显性单基因遗传.例如, 王新华(2010)通过构建F2群体对TuMV-C5进行抗性鉴定及彭海涛等(2012b)通过构建F2和BC1群体对TuMV-C4进行抗性鉴定, 均表明不结球白菜对TuMV的抗性符合单基因显性模型. ... 大白菜BrCNGC全基因组鉴定及其表达分析 1 2018 ... 除了通过分子标记鉴定TuMV抗性相关基因外, 研究人员还通过生物信息学手段预测或进行多组学分析得到大量候选基因.基因家族是结构和功能上具有相似性的一组基因, 筛选鉴定与病毒相关的基因家族是快速高效获得候选基因的方法.Lv等(2015)通过生物信息学手段, 对大白菜TIR-NBS-LRR基因家族进行了全基因组鉴定, 并对部分基因在TuMV侵染下的表达模式进行分析, 最终筛选到多个与TuMV的CP基因表达模式密切相关的候选基因.SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptor)家族基因参与蛋白和膜泡的胞内及胞间运输, 与病毒侵染密切相关(Wei et al., 2013).Zhang等(2018)通过生物信息学手段对大白菜中与膜泡运输相关的SNARE基因家族进行鉴定分析, 最终得到9个可能与病毒胞间运输相关的基因, 15个可能与病毒长距离运输相关的基因.汪影等(2018)对大白菜BrCNGC (cyclic nucleotide gated ion channel)基因进行鉴定, 分别在外施脱落酸(ABA)、水杨酸(SA)和茉莉酸甲酯(MeJA)后接种TuMV, 发现多个差异表达的BrCNGCs. ... 大白菜、萝卜芜菁花叶病毒系统进化及CP序列分析 1 2018 ... 随着科技的进步, 分子生物学技术的应用使病毒株系鉴定更加精确和灵敏.研究人员使用抗体从血清学角度对TuMV进行了分类鉴定.Jenner等(1999)使用基于TuMV外壳蛋白(coat protein, CP)设计的30种单克隆抗体, 将41种病毒分离物分为3个株系.随后有学者根据基因组序列特异性对TuMV进行分类鉴定.例如, Ohshima等(2002)和Tomimura等(2003)将生物学与分子生物学技术相结合, 把TuMV分为能侵染白菜、油菜和萝卜的BR (Brassica-Raphanus)型以及不能侵染萝卜的B (Brassica)型, 这2种类型又可分为basal-B、basal-BR、Asian-BR和world-B四组.近年来, 研究人员利用此方法对山东、河南、辽宁、黑龙江及陕西等省份发病的大白菜进行分类, 发现我国各地普遍流行basal-BR组和world-B组, 且以world-B组为主(吴斌等, 2018).Sánchez等(2003)对TuMV的CP基因序列进行多态性分析, 将60个病毒分离物分成MB (mostly Brassica isolates)和MR (mostly radish isolates) 2大类, 其中MR类又可以分为IBR (in- termediate between Brassica and radish clusters)和OBR (outside Brassica and radish clusters). ... 1 2000 ... 白菜抗TuMV的遗传规律非常复杂, 国内外相关研究很多, 但结论不一.早在1980年, Provvidenti (1980)使用多个大白菜品种进行了TuMV抗性分析, 发现大白菜对TuMV具有株系特异抗性, 其抗性基因为显性遗传.Leung和Williams (1983)研究表明, TuMV-C1毒株也受显性基因控制.1984年, 我国学者钮心恪(1984)分析了抗、感亲本杂交后代F1、F2和BC1, 表明大白菜对TuMV的抗性表现为显性遗传, 由2个相互独立的基因控制.后续又有多项研究得到了相似的结论(Suh et al., 1995; 阎瑾琦, 2000; Jin et al., 2014).但Yoon等(1993)使用高抗材料0-2 (高抗TuMV-C1、C2、C3、C4和C5毒株)和3个感病材料及其F1、F2和BC1进行遗传分析, 发现了2个抗TuMV-C4和TuMV-C5毒株的隐性基因.后续也有研究得出大白菜对TuMV的抗性表现为隐性遗传的结论(韩和平, 2003; Rusholme et al., 2007; Li et al., 2011; 满卫萍, 2020).与以上2种模式不同, 鹿英杰等(1988)研究表明, 大白菜对TuMV的抗性符合加性-显性效应模型, 属于数量遗传.张俊华等(2008)和屈淑平等(2009)分别用高抗TuMV-C3株系的自交系A52-2与感病材料进行分析, 均得到了相同的结论, 即大白菜对TuMV- C3毒株的抗性为不完全显性.张明等(2011)用高抗材料A52-2进行分析, 发现大白菜对TuMV-C4的抗性也是不完全显性, 至少受5对显性基因控制.钱伟等(2012)使用ELISA方法对接种TuMV的大白菜进行鉴定, 分析了P1、P2、F1和F2四世代遗传模型, 表明TuMV-C4抗性符合2对主基因+多基因的遗传模式, 支持数量性状基因控制TuMV抗性的观点.李国亮等(2019)用高抗TuMV的89B和高感的强势为亲本构建 F2群体并进行分析, 结果也支持数量遗传模型. ... 白菜病毒诱导基因沉默技术体系的建立 1 2012 ... 综上, 白菜类作物抗TuMV相关研究取得了较大进展, 已发现多个抗性基因, 为今后分子育种和抗病毒品种的选育奠定了良好的基础.同时还可为芸薹属B、C基因组的抗病品种选育提供基因资源, 并为抗病毒分子育种提供重要参考.下一步可通过远缘杂交、体细胞融合或基因工程技术将抗性基因导入B、C基因组, 从而创制新的抗病材料.目前, 芸薹属作物特别是A基因组作物的转基因技术尚不成熟.作为替代病毒诱导的基因沉默技术已在白菜类作物中广泛应用, 可实现在不转基因的情况下快速对筛选到的候选基因进行功能验证(杨学东等, 2012; Yu et al., 2018).目前, 对于转基因蔬菜还存在一定的争议, 因此可使用CRISPR/Cas9技术, 将eIF(iso)4E等寄主的关键位点突变, 然后经过杂交等手段去除外源基因, 该技术已在拟南芥中得到成功应用(Pyott et al., 2016).同时, 还可通过杂交将多个自然群体中的抗性基因聚合到品质优良的材料中. ... TuMV-Nib反义基因对大白菜的遗传转化研究 1 2006 ... 目前, 研究人员已获得多个抗性相关的基因, 多种生物技术也应用于白菜品质改良(刘栓桃等, 2004), 将这些基因转入白菜或将其突变, 可增强TuMV抗性, 从而提高白菜的产量和品质, 并降低农药使用量.鉴于前期研究发现eIF(iso)4E的突变体对TuMV具有广谱抗性, Kim等(2014)将eIF(iso)4E (W95L、W95L/ K150E)转入大白菜感病品种, 发现可提高TuMV抗性.Nellist等(2014)发现retr01和retr02均为BraA.eIF (iso)4E.a的插入突变体, 突变后基因不能正常剪接, 因此不能被TuMV利用, 将其转入拟南芥eIF(iso)4E突变体后得到抗性植株.除以上直接转入抗性基因外, 还可以将TuMV的编码基因设计成RNAi靶标, 然后转入白菜类作物, 从而提高抗病性.例如, 朱常香等(2001)将TuMV的CP基因转入大白菜, 明显增强其抗病能力.于占东等(2006)和佟容等(2020b)将TuMV的NIb反义片段转入大白菜, 得到多个抗TuMV-C4植株.为了获得既抗TuMV又耐抽薹的大白菜, 佟容等(2020a)利用TuMV-Nib和FLY基因构建了双价反义表达载体并转化大白菜, 最终获得7株同时具有抗病毒和耐抽薹特性的植株. ... 大白菜抗芜菁花叶病毒的QTL分析 3 2008 ... 白菜抗TuMV的遗传规律非常复杂, 国内外相关研究很多, 但结论不一.早在1980年, Provvidenti (1980)使用多个大白菜品种进行了TuMV抗性分析, 发现大白菜对TuMV具有株系特异抗性, 其抗性基因为显性遗传.Leung和Williams (1983)研究表明, TuMV-C1毒株也受显性基因控制.1984年, 我国学者钮心恪(1984)分析了抗、感亲本杂交后代F1、F2和BC1, 表明大白菜对TuMV的抗性表现为显性遗传, 由2个相互独立的基因控制.后续又有多项研究得到了相似的结论(Suh et al., 1995; 阎瑾琦, 2000; Jin et al., 2014).但Yoon等(1993)使用高抗材料0-2 (高抗TuMV-C1、C2、C3、C4和C5毒株)和3个感病材料及其F1、F2和BC1进行遗传分析, 发现了2个抗TuMV-C4和TuMV-C5毒株的隐性基因.后续也有研究得出大白菜对TuMV的抗性表现为隐性遗传的结论(韩和平, 2003; Rusholme et al., 2007; Li et al., 2011; 满卫萍, 2020).与以上2种模式不同, 鹿英杰等(1988)研究表明, 大白菜对TuMV的抗性符合加性-显性效应模型, 属于数量遗传.张俊华等(2008)和屈淑平等(2009)分别用高抗TuMV-C3株系的自交系A52-2与感病材料进行分析, 均得到了相同的结论, 即大白菜对TuMV- C3毒株的抗性为不完全显性.张明等(2011)用高抗材料A52-2进行分析, 发现大白菜对TuMV-C4的抗性也是不完全显性, 至少受5对显性基因控制.钱伟等(2012)使用ELISA方法对接种TuMV的大白菜进行鉴定, 分析了P1、P2、F1和F2四世代遗传模型, 表明TuMV-C4抗性符合2对主基因+多基因的遗传模式, 支持数量性状基因控制TuMV抗性的观点.李国亮等(2019)用高抗TuMV的89B和高感的强势为亲本构建 F2群体并进行分析, 结果也支持数量遗传模型. ...
黑色粗体表示通过分子标记筛选到的TuMV抗性相关基因或位点.黑色非粗体表示通过同源克隆、基因家族分析或组学分析筛选得到的候选基因.实心黑点和实心正方形分别表示通过对TIR-NBS-LRR家族和SNARE家族分析得到的候选基因; 实心正三角、实心倒三角形和实心菱形分别表示通过蛋白质组分析、全基因组关联分析和代谢组分析得到的候选基因.为便于分析, 根据最新注释将定位于Scaffold000104上的Bra035393定位到A04染色体的对应位置.本图基于Brara_Chiifu_V1.5基因组(Wang et al., 2011b)使用MG2C v2.1软件(Chao et al., 2021)绘制. ... Genome-wide identification of turnip mosaic virus-responsive microRNAs in non-heading Chinese cabbage by high-throughput sequencing 1 2015 ... 与基因家族分析相比, 组学分析可以获得更高通量且更全面的数据.Wang等(2015)通过对接种TuMV的不结球白菜进行microRNA测序, 得到45个新的microRNA及69个差异表达microRNA, 并发现部分microRNA同时响应TuMV侵染和低温处理.Liu等(2019)使用同位素标记相对和绝对定量(iTRAQ)技术, 对接种TuMV后第7、14和21天不结球白菜的茎和叶柄进行蛋白质组学分析, 发现TuMV的长距离运输与多个调控途径相关, 在差异表达蛋白中有多个与热激蛋白、病程相关蛋白、活性氧清除蛋白、胞间连丝和钙离子转运相关.Yu等(2019)对接种TuMV的大白菜进行全基因组DNA甲基化分析, 发现差异甲基化基因中与抗病相关的基因主要集中在NBS基因家族、生长素与SA信号转导、细胞壁合成及蛋白质降解等途径, 推测DNA甲基化主要通过调节这些基因来激活植物的免疫反应.Zhang等(2020)通过全基因组关联分析(genome wide association study, GWAS), 对接种TuMV的83个不结球白菜株系进行分析, 共鉴定到17个相关位点, 随后选取9个基因进行验证, 对相应的拟南芥同源基因突变体接种TuMV, 结果发现细胞壁相关蛋白(cell wall-associated proteins)、PME (pectin methyl-esterase)、TFs (transcription factors)、R (resistance)、VAN3/SFC protein和F-box相关基因的突变体中TuMV含量显著高于对照, 表明这些基因可能是抗病基因.为了得到更全面可靠的数据, 我们对接种TuMV的大白菜进行转录组和蛋白质组联合分析, 在转录组和蛋白质组中分别得到多个可能与TuMV侵染相关的候选基因, 还得到12个相关性很高的基因和蛋白, 其中包括差异表达最显著的非特异脂质转运蛋白nsLTP (nonspecific lipid transfer protein) (吕善武, 2019; Lyu et al., 2020).已有研究显示脂质转运蛋白可以提高植物的抗病毒能力(Park et al., 2002; Sarowar et al., 2009).我们认为大白菜nsLTPs很可能是抗病基因, 但其抗性机理还有待阐明.最近, Lu等(2022)对挥发性物质进行代谢组与转录组联合分析, 结果表明, TuMV可能导致挥发性有机化合物含量增加, 芸薹属特有的吸引蚜虫的物质异硫氰酸烯丙基在白菜感染TuMV三周后消失. ... The SNARE protein Syp71 is essential for turnip mo- saic virus infection by mediating fusion of virus-induced vesicles with chloroplasts 1 2013 ... 除了通过分子标记鉴定TuMV抗性相关基因外, 研究人员还通过生物信息学手段预测或进行多组学分析得到大量候选基因.基因家族是结构和功能上具有相似性的一组基因, 筛选鉴定与病毒相关的基因家族是快速高效获得候选基因的方法.Lv等(2015)通过生物信息学手段, 对大白菜TIR-NBS-LRR基因家族进行了全基因组鉴定, 并对部分基因在TuMV侵染下的表达模式进行分析, 最终筛选到多个与TuMV的CP基因表达模式密切相关的候选基因.SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptor)家族基因参与蛋白和膜泡的胞内及胞间运输, 与病毒侵染密切相关(Wei et al., 2013).Zhang等(2018)通过生物信息学手段对大白菜中与膜泡运输相关的SNARE基因家族进行鉴定分析, 最终得到9个可能与病毒胞间运输相关的基因, 15个可能与病毒长距离运输相关的基因.汪影等(2018)对大白菜BrCNGC (cyclic nucleotide gated ion channel)基因进行鉴定, 分别在外施脱落酸(ABA)、水杨酸(SA)和茉莉酸甲酯(MeJA)后接种TuMV, 发现多个差异表达的BrCNGCs. ... Inheritance of resistance to turnip mosaic virus in Chinese cabbage 1 1993 ... 白菜抗TuMV的遗传规律非常复杂, 国内外相关研究很多, 但结论不一.早在1980年, Provvidenti (1980)使用多个大白菜品种进行了TuMV抗性分析, 发现大白菜对TuMV具有株系特异抗性, 其抗性基因为显性遗传.Leung和Williams (1983)研究表明, TuMV-C1毒株也受显性基因控制.1984年, 我国学者钮心恪(1984)分析了抗、感亲本杂交后代F1、F2和BC1, 表明大白菜对TuMV的抗性表现为显性遗传, 由2个相互独立的基因控制.后续又有多项研究得到了相似的结论(Suh et al., 1995; 阎瑾琦, 2000; Jin et al., 2014).但Yoon等(1993)使用高抗材料0-2 (高抗TuMV-C1、C2、C3、C4和C5毒株)和3个感病材料及其F1、F2和BC1进行遗传分析, 发现了2个抗TuMV-C4和TuMV-C5毒株的隐性基因.后续也有研究得出大白菜对TuMV的抗性表现为隐性遗传的结论(韩和平, 2003; Rusholme et al., 2007; Li et al., 2011; 满卫萍, 2020).与以上2种模式不同, 鹿英杰等(1988)研究表明, 大白菜对TuMV的抗性符合加性-显性效应模型, 属于数量遗传.张俊华等(2008)和屈淑平等(2009)分别用高抗TuMV-C3株系的自交系A52-2与感病材料进行分析, 均得到了相同的结论, 即大白菜对TuMV- C3毒株的抗性为不完全显性.张明等(2011)用高抗材料A52-2进行分析, 发现大白菜对TuMV-C4的抗性也是不完全显性, 至少受5对显性基因控制.钱伟等(2012)使用ELISA方法对接种TuMV的大白菜进行鉴定, 分析了P1、P2、F1和F2四世代遗传模型, 表明TuMV-C4抗性符合2对主基因+多基因的遗传模式, 支持数量性状基因控制TuMV抗性的观点.李国亮等(2019)用高抗TuMV的89B和高感的强势为亲本构建 F2群体并进行分析, 结果也支持数量遗传模型. ... On the strain distribution of turnip mosaic virus 1 1963 ... 1963年, Yoshii (1963)根据侵染心叶烟(Nicotiana glutinosa)和甘蓝后产生症状的严重程度, 将TuMV分为普通株系(ordinary strain)和甘蓝株系(cabbage strain).McDonald和Hiebert (1974)根据宿主范围将TuMV分为2种类型, I型可侵染所有芸薹属植物, II型只能侵染部分芸薹属植物. ... The methylation pattern of DNA and complex correlations with gene expressions during TuMV infection in Chinese cabbage 1 2019 ... 与基因家族分析相比, 组学分析可以获得更高通量且更全面的数据.Wang等(2015)通过对接种TuMV的不结球白菜进行microRNA测序, 得到45个新的microRNA及69个差异表达microRNA, 并发现部分microRNA同时响应TuMV侵染和低温处理.Liu等(2019)使用同位素标记相对和绝对定量(iTRAQ)技术, 对接种TuMV后第7、14和21天不结球白菜的茎和叶柄进行蛋白质组学分析, 发现TuMV的长距离运输与多个调控途径相关, 在差异表达蛋白中有多个与热激蛋白、病程相关蛋白、活性氧清除蛋白、胞间连丝和钙离子转运相关.Yu等(2019)对接种TuMV的大白菜进行全基因组DNA甲基化分析, 发现差异甲基化基因中与抗病相关的基因主要集中在NBS基因家族、生长素与SA信号转导、细胞壁合成及蛋白质降解等途径, 推测DNA甲基化主要通过调节这些基因来激活植物的免疫反应.Zhang等(2020)通过全基因组关联分析(genome wide association study, GWAS), 对接种TuMV的83个不结球白菜株系进行分析, 共鉴定到17个相关位点, 随后选取9个基因进行验证, 对相应的拟南芥同源基因突变体接种TuMV, 结果发现细胞壁相关蛋白(cell wall-associated proteins)、PME (pectin methyl-esterase)、TFs (transcription factors)、R (resistance)、VAN3/SFC protein和F-box相关基因的突变体中TuMV含量显著高于对照, 表明这些基因可能是抗病基因.为了得到更全面可靠的数据, 我们对接种TuMV的大白菜进行转录组和蛋白质组联合分析, 在转录组和蛋白质组中分别得到多个可能与TuMV侵染相关的候选基因, 还得到12个相关性很高的基因和蛋白, 其中包括差异表达最显著的非特异脂质转运蛋白nsLTP (nonspecific lipid transfer protein) (吕善武, 2019; Lyu et al., 2020).已有研究显示脂质转运蛋白可以提高植物的抗病毒能力(Park et al., 2002; Sarowar et al., 2009).我们认为大白菜nsLTPs很可能是抗病基因, 但其抗性机理还有待阐明.最近, Lu等(2022)对挥发性物质进行代谢组与转录组联合分析, 结果表明, TuMV可能导致挥发性有机化合物含量增加, 芸薹属特有的吸引蚜虫的物质异硫氰酸烯丙基在白菜感染TuMV三周后消失. ... Efficient virus-induced gene silencing in Brassica rapa using a turnip yellow mosaic virus vector 1 2018 ... 综上, 白菜类作物抗TuMV相关研究取得了较大进展, 已发现多个抗性基因, 为今后分子育种和抗病毒品种的选育奠定了良好的基础.同时还可为芸薹属B、C基因组的抗病品种选育提供基因资源, 并为抗病毒分子育种提供重要参考.下一步可通过远缘杂交、体细胞融合或基因工程技术将抗性基因导入B、C基因组, 从而创制新的抗病材料.目前, 芸薹属作物特别是A基因组作物的转基因技术尚不成熟.作为替代病毒诱导的基因沉默技术已在白菜类作物中广泛应用, 可实现在不转基因的情况下快速对筛选到的候选基因进行功能验证(杨学东等, 2012; Yu et al., 2018).目前, 对于转基因蔬菜还存在一定的争议, 因此可使用CRISPR/Cas9技术, 将eIF(iso)4E等寄主的关键位点突变, 然后经过杂交等手段去除外源基因, 该技术已在拟南芥中得到成功应用(Pyott et al., 2016).同时, 还可通过杂交将多个自然群体中的抗性基因聚合到品质优良的材料中. ... Genome-wide identification, classification, and expression analysis of SNARE genes in Chinese cabbage (Brassica rapa ssp. pekinensis) infected by turnip mosaic virus 1 2018 ... 除了通过分子标记鉴定TuMV抗性相关基因外, 研究人员还通过生物信息学手段预测或进行多组学分析得到大量候选基因.基因家族是结构和功能上具有相似性的一组基因, 筛选鉴定与病毒相关的基因家族是快速高效获得候选基因的方法.Lv等(2015)通过生物信息学手段, 对大白菜TIR-NBS-LRR基因家族进行了全基因组鉴定, 并对部分基因在TuMV侵染下的表达模式进行分析, 最终筛选到多个与TuMV的CP基因表达模式密切相关的候选基因.SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptor)家族基因参与蛋白和膜泡的胞内及胞间运输, 与病毒侵染密切相关(Wei et al., 2013).Zhang等(2018)通过生物信息学手段对大白菜中与膜泡运输相关的SNARE基因家族进行鉴定分析, 最终得到9个可能与病毒胞间运输相关的基因, 15个可能与病毒长距离运输相关的基因.汪影等(2018)对大白菜BrCNGC (cyclic nucleotide gated ion channel)基因进行鉴定, 分别在外施脱落酸(ABA)、水杨酸(SA)和茉莉酸甲酯(MeJA)后接种TuMV, 发现多个差异表达的BrCNGCs. ... Quantitative trait loci analysis for resistance against turnip mosaic virus based on a doubled-haploid population in Chinese cabbage 2 2008 ... The TuMV resistance genes and makers in Brassica rapa
本文的其它图/表
|

 首页
首页