
|
|
||
|
植物ESCRT复合体的功能研究进展
植物学报
2022, 57 (5):
697-712.
DOI: 10.11983/CBB22038
真核细胞中, 功能高度保守的内体蛋白分选转运装置ESCRT在胞吞途径和蛋白分泌途径中均扮演重要角色。植物细胞中, 该装置包含ESCRT-I、ESCRT-II、ESCRT-III和VPS4/SKD1复合体4个亚基, 但缺乏ESCRT-0亚基。ESCRT的每个亚基均由多个蛋白构成。目前, 针对ESCRT的研究已经证实, 其在泛素化的膜蛋白进入多囊泡体/液泡前体(MVB/PVC)内腔过程中发挥重要调控作用; 同时在自噬途径以及应对环境胁迫等方面也具有重要的调节功能。该文首先介绍了植物中ESCRT复合体的组成及生物学功能, 然后总结了植物中特有ESCRT复合体组分蛋白的最新研究进展, 最后探讨了有关ESCRT复合体研究中尚未解决的重要科学问题。  View image in article
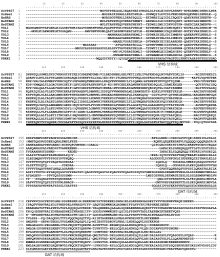
图4
不同物种中ESCRT-III组分蛋白(VPS2 (A)、VPS20 (B)、VPS24 (C)和SNF7 (D))间的氨基酸序列比对
横线区域分别表示保守的CC结构域和SNF7结构域。
正文中引用本图/表的段落
在酵母和动物细胞中, ESCRT-III由4个亚基组成, 即VPS2p/CHMP2 (charged multivesicular body protein 2)、VPS20p/CHMP6、VPS24p/CHMP3和SNF7p (sucrose non-fermenting 7)/CHMP4。此外, 还包括Did2 (Doa4-independent degradation2)/CHMP1、VPS60/CHMP5和IST1 (increased salt tolerance 1) 3个辅助蛋白(Teis et al., 2008)。在内吞途径中, ESCRT-III组分蛋白VPS20首先与ESCRT-II组分蛋白VPS25互作而激活ESCRT-III, 同时SNF7多聚体被VPS20招募到MVB/PVC膜上组装ESCRT-III, 并进一步招募VPS2和VPS24等蛋白到MVB/PVC膜形成成熟的ESCRT-III复合体, 从而介导膜的凹陷和剪切形成ILVs。Did2/CHMP1、VPS60/CHMP5和IST1三个辅助蛋白则通过激活VPS4/SKD1-LIP5复合体活性参与调节ESCRT-III与VPS4/SKD1-LIP5复合体结合的强弱、ESCRT-III从MVB/PVC膜上的解离及解聚过程, 以此调控ESCRT-III组分蛋白的循环利用和MVB/PVC的成熟(Henne et al., 2011)。拟南芥中的ESCRT-III具有高度保守性, 对应的同源蛋白均含有多个亚基, 包括VPS2.1、VPS2.2和VPS2.3, VPS20.1和VPS20.2, SNF7A和SNF7B, VPS24-1和VPS24-2 (Gao et al., 2017)以及能够与SNF7发生互作的ALIX (图4A-D)。通过酵母双杂交筛选, 发现ESCRT-III组分蛋白VPS2.1和VPS24.1均可与去泛素化酶AMSH3 (associated molecular with SH3 domain of STAM 3)互作(Katsiarimpa et al., 2011), 它们之间的互作分别依赖AMSH3的MIT (microtubule interacting and transport)结构域和ESCRT-III的MIM1 (microtubule interacting and transport (MIT)-interacting motif1)结构域。亚细胞定位研究显示, AMSH3与ESCRT-III组分蛋白共定位于E类晚期胞内吞小体(class E compartments)中, AMSH3的DUB (deubiquitinating enzyme)结构域缺失会导致ESCRT-III亚细胞定位错乱, 因此AMSH3对ESCRT-III正确的亚细胞定位具有重要作用(Katsiarimpa et al., 2011)。AMSH3定位到晚期内吞体则受到其互作蛋白ALIX的调控, ALIX基因缺失突变不仅会导致植株胚死亡, MVB/PVC和液泡形态异常, 而且会使AMSH3定位到晚期内吞体的能力消失(Kalinowska et al., 2015)。由此可见, AMSH3与ESCRT-III的协同作用依赖ALIX的功能。对AMSH3基因功能的进一步研究显示, 缺失AMSH3基因也会引起类似alix突变体的表型, 并进一步表现出泛素化蛋白和自噬体异常积累等表型(Isono et al., 2010)。此外, 对vps2.1突变体的研究发现, 突变体植株表现出胚死亡, 具有类似于chmp1a/chmp1b双突变体的表型(Spitzer et al., 2009; Katsiarimpa et al., 2011), 因此ESCRT-III复合体组分蛋白的缺失均可导致植物发育受阻而死亡。近期研究发现, 另一个去泛素化酶AMSH1也可识别并结合VPS2.1, 参与调节货物蛋白的去泛素化过程。过表达VPS2.1或缺失AMSH1基因均会导致植物出现矮化和早衰表型, 并表现出PIN2异常积累于液泡膜及其内吞降解过程受到强烈抑制, 自噬体标记物ATG8e异常积累以及自噬途径受阻等现象(Katsiarimpa et al., 2011, 2013; Schwihla and Korbei, 2020), 表明VPS2.1和2个去泛素化酶AMSH3与AMSH1在植物生长及物质循环利用过程中起重要作用。在拟南芥悬浮细胞中, 过表达活性缺失形式的vps24.1-1-152会导致MVB/PVC内腔中ILV数量显著减少, MVB/PVC形态异常(Cai et al., 2014), 同时导致生长素生物合成关键酶YUC1黄素单氧合酶(YUC1 flavin monooxygenase)无法正常进入液泡降解(Ge et al., 2019)。进一步研究发现, ESCRT-III辅助蛋白CHMP1可与VPS4/SKD1结合, 调节MVB/ PVC的形成及ESCRT-III复合体的解聚。chmp1a/ chmp1b双突变体植株中, 生长素转运载体蛋白PIN1、PIN2和AUX1在细胞膜上积累, 无法正常内吞降解, 依赖于自噬途径降解的质体(plastid)蛋白大量积累于细胞质; 并表现出胚胎或早期幼苗致死表型(Spitzer et al., 2009, 2015)。因此, ESCRT-III组分蛋白对植物是否能够存活具有重要作用; 同时, ESCRT- III组分蛋白的缺失会导致自噬体积累, 进而使依赖于自噬体途径降解的蛋白异常积累。但截至目前, 人们对ESCRT-III和自噬体协同作用的调控因子仍知之甚少, 有待进一步探究。
本文的其它图/表
|
 首页
首页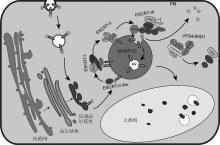

{kind=link}